Why The Heart Almost Never Gets Cancer: Mechanical Load, Nesprin-2, And The Epigenetics Of A Beating Organ
By Jacob Gordon, INHC, FMT-CThis article contains affiliate links. As an Amazon Associate, MyBioHack earns from qualifying purchases at no extra cost to you. We only link products we research and stand behind.
The heart is one of the only organs in the body that almost never develops cancer, and a 2025 Science paper finally offers a mechanistic reason why.
In this post, we will discuss why the beating heart resists tumors, how physical force gets translated into epigenetic silencing through Nesprin-2, what human cardiac metastases reveal when that protection fails, and what this says about the larger relationship between mechanical load and gene expression.

The Organ Cancer Skips
Primary tumors that start in the heart are vanishingly rare, with autopsy series reporting rates between roughly 0.001% and 0.03%. R
Three quarters of the primary tumors that do form are benign, and nearly half of those are myxomas. R
Cancer reaching the heart from elsewhere is more common than a tumor starting there, but it is still uncommon relative to how much blood the heart handles. R
This is the part that should bother you if you think about it mechanistically.
The myocardium is one of the most densely vascularized tissues in the body, so cancer cells circulating in the blood pass through it constantly.
By the logic of seeding, the heart should be a frequent metastatic site.
Instead it is one of the rarest, and the question the new work set out to answer is what, specifically, the cardiac microenvironment does to a cancer cell that other tissues do not.
What Makes The Heart Resistant
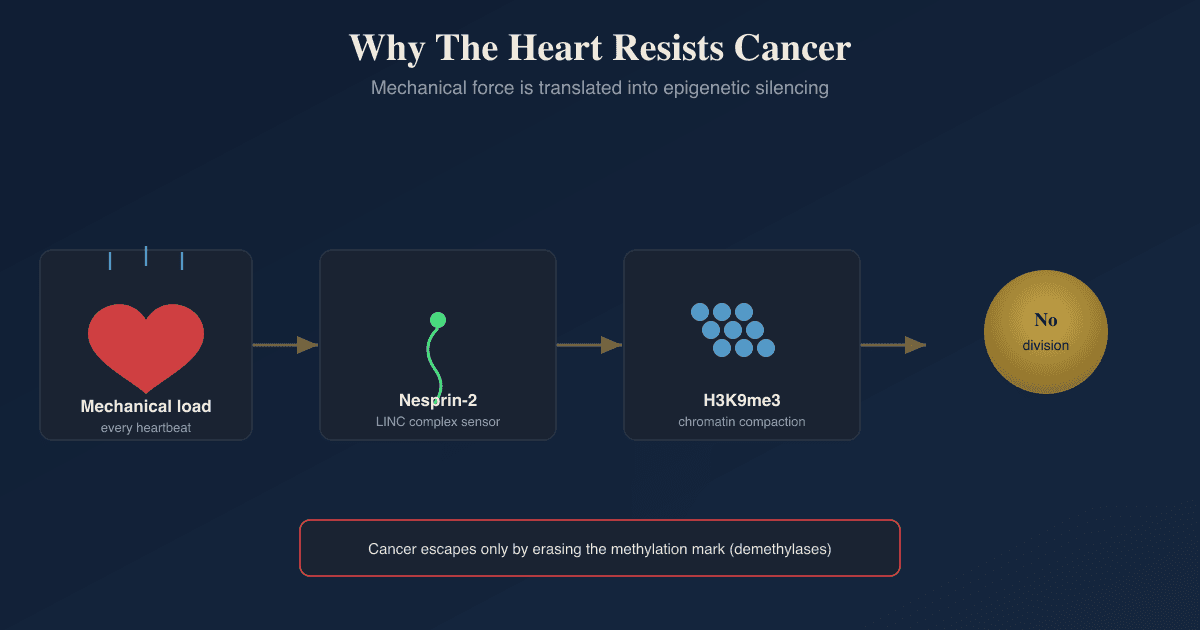
The candidate explanation the authors tested is mechanical load, meaning the constant rhythmic stretch and contraction the heart muscle generates with every beat.
This was not a random guess.
Mechanical load is already the leading explanation for why the adult heart cannot regenerate.
Cardiomyocytes proliferate freely before birth, then exit the cell cycle within the first days to weeks of life, and the rise in hemodynamic workload after birth is one of the signals that shuts that proliferation down. R
The clinical mirror image is striking.
When a failing human heart is mechanically unloaded with a left ventricular assist device, cardiomyocytes partially re-enter the cell cycle and begin proliferating again. R
So the hypothesis was simple.
If mechanical load forces the heart's own cells to stop dividing, it might do the same thing to a cancer cell that lands there. R
The authors tested this several ways.
They built a genetic mouse model in which the same oncogenic hit (mutant K-Ras plus loss of p53) was triggered across liver, skeletal muscle, and heart.
Tumors formed at multiple sites but never in the heart, despite a comparable degree of the underlying genetic recombination in cardiac tissue. R
They then used a heterotopic heart transplant model that keeps a heart perfused but mechanically unloaded, plus engineered heart tissues where load can be dialed up or down at will.
Across lung adenocarcinoma, colon carcinoma, and melanoma cells, mechanical load suppressed proliferation inside the myocardium, and removing the load let those same cells proliferate. R
That is the core finding.
The protection is not chemical and it is not immune.
It is physical, and it can be switched off by taking the load away.

How Mechanical Force Reaches The Genome
A mechanical force at the cell surface only changes a cell's behavior if something carries that force inward and converts it into a molecular signal.
That conversion is called mechanotransduction, and the structure that does it here is the Linker of Nucleoskeleton and Cytoskeleton (LINC) complex.
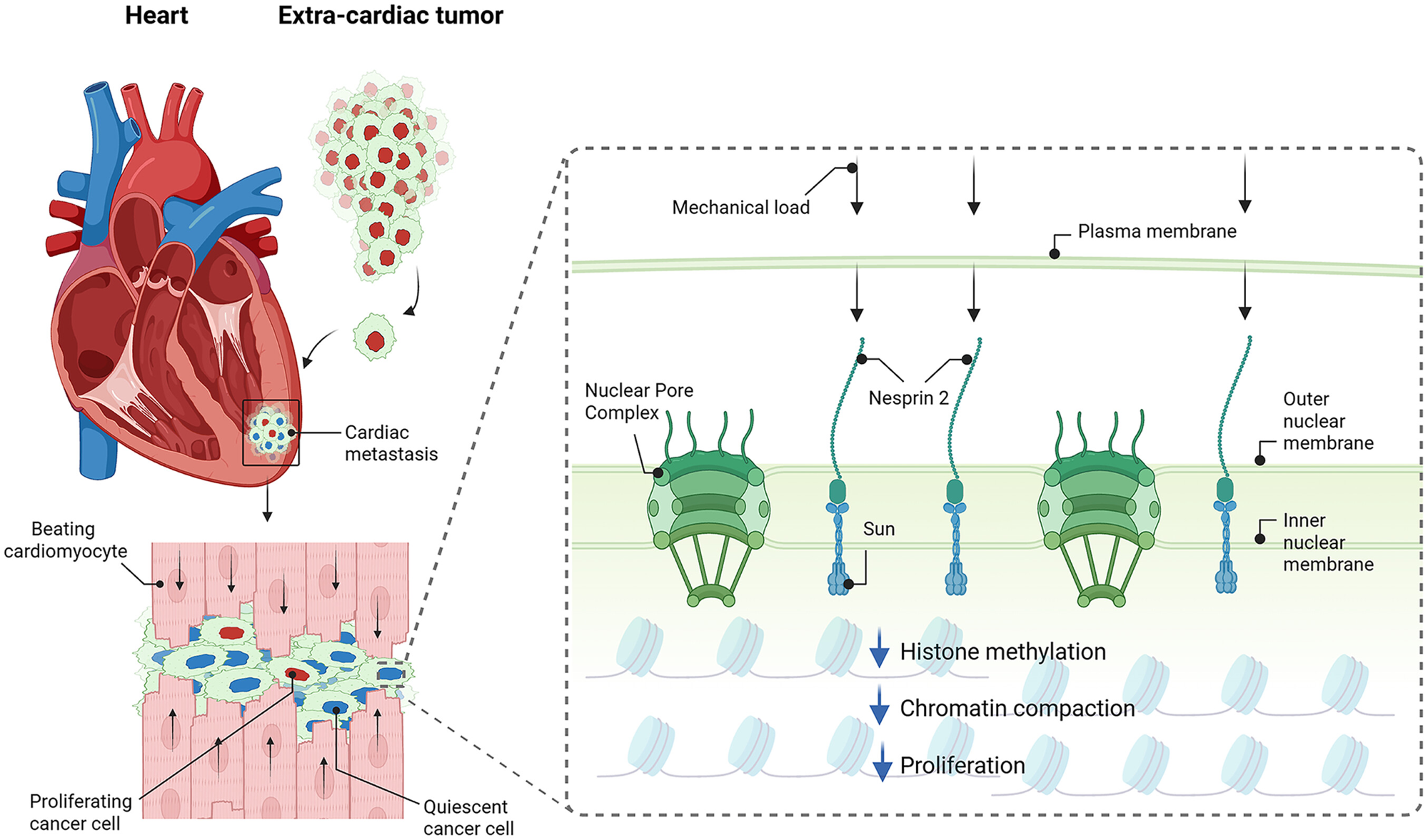
The LINC complex physically bridges the cytoskeleton on the outside of the nucleus to the chromatin on the inside, so a pull on the cell can become a pull on the genome. R
The outward-facing half of that bridge is a protein called Nesprin-2.
Nesprin-2 sits in the outer nuclear membrane, binds the actin cytoskeleton on one end, and connects through SUN-domain proteins in the inner nuclear membrane to the nuclear interior on the other end.
Force sensors built into Nesprin-2 have shown directly that it is under mechanical tension that depends on actomyosin contractility, which makes it a genuine force-bearing element rather than a passive tether. R
In the new study, Nesprin-2 emerged as the molecule that senses the beating heart's mechanical forces and translates them into reduced cancer cell proliferation. R
The cleanest piece of evidence is the loss-of-function experiment.
When the authors silenced Nesprin-2 in lung cancer cells before implanting them in the heart, the cells regained the ability to proliferate even under normal physiological load, and they formed large tumors. R
In other words, knock out the mechanical sensor and the heart loses its resistance.
The force is still there, but the cell can no longer feel it.
The Epigenetic Switch: Histone Methylation And Chromatin Compaction
This is where the story becomes an epigenetics story rather than a mechanics story.
When Nesprin-2 transmits load into the nucleus, the downstream effect is on chromatin, the way DNA is physically packaged around histone proteins.
Two changes happen together under load.
Histone methylation rises, specifically the repressive mark trimethylation of histone 3 at lysine 9 (H3K9me3), and the chromatin becomes more compacted. R
H3K9me3 is the defining chemical signature of constitutive heterochromatin, the densely packed, transcriptionally silent fraction of the genome. R
Compacted, H3K9me3-marked chromatin is also what gives the nucleus its mechanical stiffness and holds proliferation-driving genes shut. R
So the logic of the pathway is a clean chain.
Mechanical load pulls on Nesprin-2, Nesprin-2 transmits that force to the genome, the genome responds by adding H3K9me3 and compacting, and the genes a cancer cell needs to divide get locked away.
The authors confirmed this was happening at the right places, showing that chromatin accessibility and histone methylation shifted specifically at sites that control cancer cell proliferation. R
This is worth connecting to a distinction that confuses a lot of people.
The methylation being discussed here is histone methylation, a tag on the proteins DNA wraps around, which is not the same as DNA methylation, the tag placed directly on cytosine bases that we cover in the methylation guide.
Both are epigenetic, both silence genes, but they are different chemistry on different molecules, and this heart mechanism runs through the histone side.
The Human Evidence: What Cardiac Metastases Reveal
The mouse and engineered-tissue work shows what happens when you add or remove load.
The human work shows what the failure mode looks like, by asking what is different about the rare cancers that do colonize the heart.
The authors used spatial transcriptomics on human samples from patients whose cancer had seeded both a cardiac metastasis and an extracardiac one, so the same primary tumor could be compared in two locations.
The cardiac metastases shared a common transcriptional profile regardless of where the cancer started. R
Among the most up-regulated genes in those cardiac metastases were histone demethylases, the enzymes that erase methylation marks. R
That fits the mechanism exactly.
If load suppresses cancer by adding H3K9me3, then a cancer that survives in the heart should be one that has found a way to strip H3K9me3 back off, and overexpressing demethylases is precisely how a cell would do that.
Consistent with that, the cardiac metastases showed reduced H3K9me3 and reduced chromatin compaction, the opposite of what load normally enforces. R
This is the same enzymology that shows up in aggressive cancers elsewhere.
The KDM4 (JMJD2) family of H3K9 demethylases is overexpressed across many tumor types, and that overexpression drives proliferation, invasion, and anchorage-independent growth. R R
The heart story is a tissue-specific version of a general truth, which is that erasing repressive histone methylation tends to unlock a cancer cell, and maintaining it tends to restrain one. R
Mechanical Load Beyond The Heart
The reason this paper matters past cardiology is that it is one more line of evidence that physical force is an upstream regulator of gene expression, not a passive background condition.
The best-studied version of this idea is the YAP/TAZ system, the mechanosensitive arm of the Hippo pathway.
YAP and TAZ read out matrix stiffness, stretch, and cytoskeletal tension, and depending on the mechanical context they move into the nucleus and switch proliferation genes on or off. R
In stiff or mechanically deranged tissue, YAP/TAZ activation is a recognized driver of tumor proliferation and treatment resistance. R
The heart finding adds a different flavor of the same principle.
Here the mechanical signal is suppressive rather than activating, and it runs through the LINC complex and histone methylation rather than through YAP/TAZ, which tells you mechanotransduction is not one pathway with one direction.
It is a family of force-reading systems whose output depends on the tissue, the type of force, and which sensor is wired to the genome.
For a chronic illness audience this is the same conceptual thread that runs through vascular and connective-tissue biology, where mechanical properties of the matrix and the cell shape downstream signaling, and it overlaps with how mechanical and metabolic stress feed into the broader cancer-versus-degeneration tradeoff we discuss in cancer and Alzheimer's disease as opposites.
What This Does And Does Not Mean For You
This is the section where honesty matters more than excitement.
What the study establishes is a mechanism, in mice, engineered tissue, and a small set of human samples, for why the heart resists cancer.
What it does not establish is a treatment, a supplement, or a protocol.
There is a big MAYBE sitting between this biology and anything actionable, and it would be dishonest to skip over it.
A few things are worth saying clearly.
The protective force here is a very specific kind, the rhythmic high-frequency load of cardiac muscle, and it is not the same as lifting weights or doing cardio, so this is not evidence that exercise compresses your chromatin to fight tumors.
Exercise has real and well-documented benefits, and we cover the molecular side of that in exercise mimetics, but extrapolating this exact heart mechanism to a gym session is not supported by the data.
The translational hope the authors actually raise is the reverse direction, meaning therapies that deliver controlled mechanical stimulation, or that target Nesprin-2 signaling and histone demethylases, to push cancer cells toward the quiescent state the heart naturally enforces. R
That is a research direction, not a recommendation.
The honest takeaway is conceptual.
Your genome is not only listening to what you eat, what you breathe, and what your hormones are doing.
It is also listening to physical force, and in at least one tissue that force is strong enough to hold cancer in check on its own.
Compounds, Herbs, And Drugs That Touch This Pathway
Start with the honest headline.
There is no supplement, herb, or drug that has been shown to directly and safely raise Nesprin-2, and anyone selling you one is guessing.
The pharmacology of SYNE2 (the gene encoding Nesprin-2) and the wider LINC complex is mostly the pharmacology of breaking it on purpose to study it, using tools like the actin-disrupting agent latrunculin A or dominant-negative constructs, which is the opposite of what you would want. R
So the realistic levers are not the sensor itself.
They sit one step upstream, at the methyl donors that let the protective mark get written, and one step downstream, at the enzymes that erase it, plus the mechanical input the study actually validated.
I am grading every item here honestly, because almost none of it has been tested for this purpose in humans.
The Mechanical Lever
The only intervention the study itself points to is mechanical, since the protective signal is physical force, and the authors frame the translational hope as controlled mechanical stimulation rather than a drug. R
The conceptually closest thing you can do is load tissue mechanically through resistance training, but I will repeat the caveat from above, which is that the rhythmic high-frequency load of heart muscle is not the same as lifting weights, and no human data shows exercise reproduces this specific chromatin compaction.
We cover the molecular side of mechanical and metabolic loading in exercise mimetics.
Keeping The Protective Mark On (Methyl-Donor Support)
Every histone methylation reaction, including the H3K9me3 that load uses to silence proliferation, runs on the same methyl donor, S-adenosylmethionine (SAM), produced by the one-carbon and methylation cycle. R
H3K9 trimethylation is one of the marks most sensitive to a drop in SAM, so methyl-donor adequacy is permissive for this pathway even though it does not specifically aim at it. R
This is the most defensible and least exotic lever, and it is the same machinery covered in the methylation guide.
The supplements that support methylation capacity: (not an exclusive list)
- Betaine (TMG) (feeds the BHMT route that regenerates methionine)
- Methylcobalamin (cofactor for methionine synthase)
- Methylfolate (5-MTHF) (carries the one-carbon unit into the cycle)
- Riboflavin (B2) (cofactor for MTHFR)
- S-Adenosylmethionine (SAMe) (the methyl donor itself)
The honest caveat is that flooding the methylation cycle supports the writing machinery in general, it does not point it at H3K9 in particular, so this removes a constraint rather than driving the heart mechanism.
Restraining The Erasers (Histone Demethylase Inhibition)
Because the cancers that colonize the heart escape by overexpressing H3K9 demethylases, the mechanistically aligned lever is to restrain those demethylases, mainly the KDM4 (JMJD2) family. R
A number of dietary flavonoids and catechols have been identified as Jumonji-C demethylase inhibitors in screening and docking studies, including the flavone orientin against KDM4C and scaffolds derived from the natural product purpurogallin. R R
The caveats here are large and I will not soften them.
These are in vitro and in silico findings rather than human outcomes, the concentration that inhibits a demethylase in a dish is not what a flavonoid supplement delivers to a nucleus, and KDM4 has normal physiological roles, so blanket demethylase inhibition is not automatically good.
This is a big MAYBE, and the right reading is mechanistic interest, not a protocol.
The Vitamin C Landmine
This is the counterintuitive part that most people get backward.
The H3K9 demethylases are Fe(II) and 2-oxoglutarate dependent dioxygenases that use vitamin C as a cofactor, so high-dose ascorbate enhances their activity. R R
Within this specific mechanism that means vitamin C pushes H3K9me3 down, the opposite direction from the heart's protection, which is the reverse of the assumption that antioxidants are always protective.
This is not a reason to avoid vitamin C, whose systemic benefits are real and large, but it is a reason not to assume that piling on antioxidants reinforces this particular pathway.
Drugs (Research-Only)
Selective KDM4 small-molecule inhibitors are in preclinical development as anticancer agents, and they are the cleanest pharmacological version of keeping H3K9me3 switched on, but none are approved or available. R
Chaetocin, a natural product from Chaetomium fungi, is the compound most often described as an H3K9 methyltransferase (SUV39H1) inhibitor and has anticancer activity in leukemia and glioma models, but note that it lowers H3K9 methylation, the opposite of the heart's protective direction, which is a useful reminder that the same mark can be pro- or anti-tumor depending on context. R
Testing
There is no consumer lab test for cardiac mechanotransduction, H3K9me3 status, or Nesprin-2 signaling, and anyone selling you one is selling you nothing.
The honest position is that this mechanism lives in research assays (single-nuclei chromatin accessibility, chromatin immunoprecipitation sequencing, spatial transcriptomics) that are not clinically available and would not change your decisions if they were.
If your underlying concern is cancer-risk biology in general rather than this specific pathway, the productive levers are the measurable upstream drivers we cover elsewhere, including chronic inflammation, metabolic dysfunction, and methylation status from the methylation guide.
For tracking those broader markers over time, the Health Hub and a consult are the right tools, rather than any test claiming to measure the heart mechanism described here.
Mechanisms Of Action
Simple:
- Every heartbeat physically squeezes and stretches the cells inside it, and that constant mechanical pressure packs their DNA so tightly that the genes a cancer cell needs to multiply cannot switch on.
- A cancer cell only escapes when it either loses the protein that feels the squeeze, or makes enzymes that pry the packed DNA back open.
Advanced:
- Force transmission through the LINC complex. Cyclic mechanical load on the cardiomyocyte is transmitted through the actin cytoskeleton to Nesprin-2 in the outer nuclear membrane, then through SUN-domain proteins spanning the perinuclear space to the inner nuclear membrane and nuclear lamina, delivering tension directly onto chromatin. R R
- Mechanically induced heterochromatin formation. Sustained nuclear tension increases H3K9me3 deposition and drives chromatin compaction into transcriptionally silent heterochromatin, which both stiffens the nucleus and represses the cell-cycle genes required for proliferation, producing a quiescent state at loci controlling cancer cell division. R R
- Escape by demethylation. Cancer cells that successfully colonize the heart overexpress histone demethylases, lowering H3K9me3 and decompacting chromatin to reopen proliferation genes, mirroring the oncogenic role of the KDM4/JMJD2 demethylase family in tumors throughout the body. R R
- Sensor-dependence. Silencing Nesprin-2 uncouples the load from the genome, so physiological mechanical force is present but no longer transduced, restoring proliferation and tumor formation in the loaded heart. R
Genetics
These are expression-level findings from tumor tissue rather than common consumer SNPs, so the genetics here is about which genes are turned up or down, not which variant you carry.
SYNE2
SYNE2 encodes Nesprin-2, the outer-nuclear-membrane protein that links the actin cytoskeleton to the nuclear envelope as part of the LINC complex.
It is the force-sensing node of this pathway, and silencing it in cancer cells removed the heart's mechanical protection and allowed tumors to grow under normal load. R
SUN1 / SUN2
SUN1 and SUN2 encode the inner-nuclear-membrane SUN-domain proteins that pair with Nesprin-2 to complete the LINC bridge across the nuclear envelope.
They carry the force from Nesprin-2 across the perinuclear space to the nuclear lamina and chromatin, making them structural partners in mechanotransduction. R
KDM4A / KDM4B / KDM4C (JMJD2 family)
The KDM4 genes encode histone demethylases that remove the repressive H3K9me2/3 marks.
They were among the up-regulated genes in human cardiac metastases, and across cancer types their overexpression strips H3K9me3, decompacts chromatin, and drives proliferation, making them the most likely escape route from this mechanism. R R
SUV39H1
SUV39H1 encodes the methyltransferase that writes H3K9me3, the opposite enzymatic activity to KDM4.
It is broadly tumor-suppressive because the heterochromatin it builds keeps proliferation genes silenced, which is the same restraint mechanical load recruits in the heart. R
More Research
A few open questions and adjacent findings add genuine value here without being settled.
Engineered heart tissue was central to this work, and the fact that load can be controlled at will in that system is what let the authors move from correlation (the heart resists cancer) to causation (load suppresses proliferation, unloading restores it), which is the methodological reason the conclusion is strong. R
Mechanotransduction is not one-directional, since the suppressive LINC-and-histone-methylation axis described here runs opposite to the proliferation-promoting YAP/TAZ axis activated by matrix stiffness, and how a given cell integrates multiple force-sensing systems at once is still unresolved. R
The cardiomyocyte parallel is the deeper biological thread, because the same mechanical load that arrests the heart's own cells after birth appears to arrest cancer cells that land there, suggesting tumor resistance is a side effect of the program that ended cardiac regeneration. R R
The therapeutic question the authors leave open is whether controlled mechanical stimulation, or pharmacological mimicry of Nesprin-2 signaling, could impose the heart's quiescence program on tumors elsewhere, which would be a fundamentally different approach from killing cancer cells with cytotoxic drugs. R
Jacob Gordon
INHC, FMT-C
Board Certified Health Coach
I spent years battling unexplained chronic illness before discovering biohacking, epigenetics, and functional medicine. Now I share that research at MyBioHack to help others find their own answers.
Book a ConsultationRelated Protocols & Supplements
Deep-dive chapters and recommended supplements for this topic
Electrolyte Complex
1 scoop/day
CoQ10
200mg/day
Magnesium Glycinate
400mg at bedtime