Methionine Restriction Vs Addition: The U-Shaped Amino Acid Behind Longevity, Cancer, And Frailty
By Jacob Gordon, INHC, FMT-CThis article contains affiliate links. As an Amazon Associate, MyBioHack earns from qualifying purchases at no extra cost to you. We only link products we research and stand behind.
Methionine is the one amino acid where the longevity literature tells you to eat less of it and the newest longevity research tells you to add it back.
In this post, we will discuss what methionine actually does in the body, the case for restricting it, the case for adding it, how a June 2026 Cell Metabolism study reconciles the two, the cancer paradox, the glycine counterbalance, who should do what, and how to test where you actually sit.

What Methionine Actually Does
Methionine is an essential sulfur-containing amino acid, which means the body cannot make it and you have to eat it.
It is also the start codon of nearly every protein you build, so it is structurally non-negotiable.
The interesting part is not the protein it builds but the signaling molecule it becomes.
Methionine is activated by methionine adenosyltransferase (MAT) into S-adenosylmethionine (SAM), which is the universal methyl donor for more than fifty methyltransferases that tag your DNA, histones, neurotransmitters, phospholipids, and creatine. R
After SAM hands off its methyl group it becomes S-adenosylhomocysteine (SAH), which is then hydrolyzed to homocysteine. R
The ratio of SAM to SAH is the cell's methylation index, and SAH is a potent brake on almost every methylation reaction, so the ratio matters more than either number alone. R
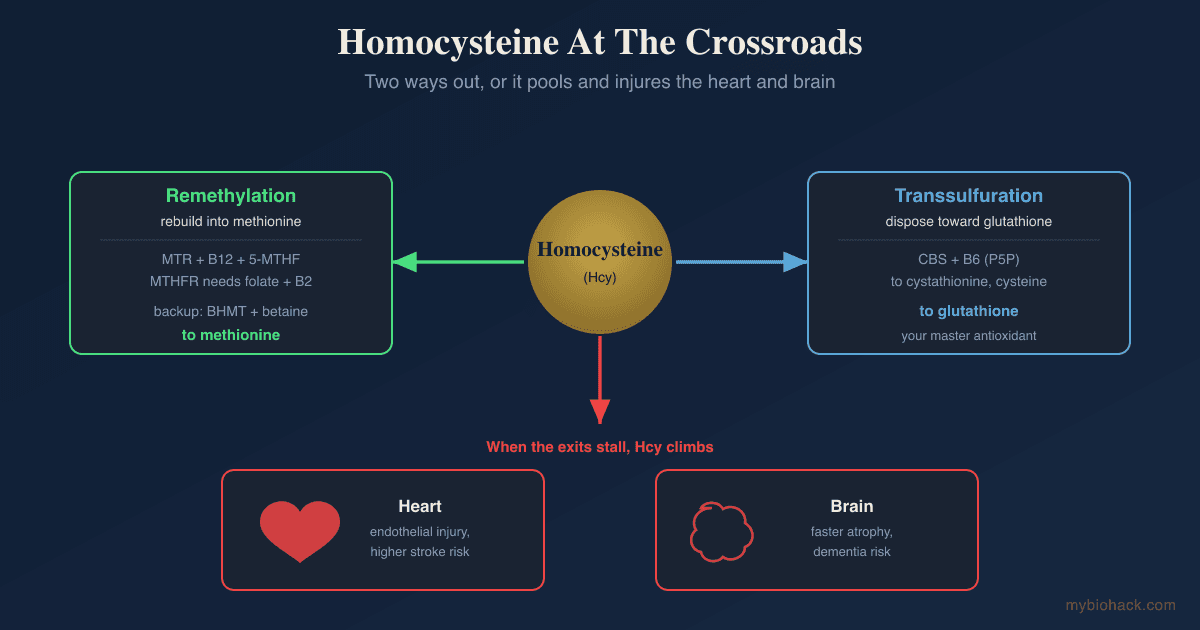
Homocysteine then has two fates, and which one it takes is the whole story.
It can be remethylated back into methionine, either by methionine synthase (MTR) using B12 and 5-methyltetrahydrofolate from the folate cycle, or by betaine-homocysteine methyltransferase (BHMT) using betaine in the liver and kidney. R
Or it can be pushed down the transsulfuration pathway by cystathionine beta-synthase (CBS), a B6-dependent enzyme that converts it toward cysteine, glutathione, and hydrogen sulfide. R

This entire web is the subject of its own deep dive in methylation: why this one pathway controls almost everything.
The reason methionine sits at the center of aging research is that SAM is also the molecule the cell uses to sense how much methionine is around.
When methionine is high, SAM is high, and a sensor called SAM sensor upstream of mTORC1 (SAMTOR) reads that and switches on the master growth pathway, mechanistic target of rapamycin complex 1 (mTORC1). R
When methionine is low, SAM drops, SAMTOR releases its grip, and mTORC1 powers down toward repair, autophagy, and stress resistance.
So methionine is not just a building block.
It is a dial that tells your cells whether to grow or to maintain.
The Case For Restriction
The restriction story is one of the most reproducible findings in the entire biology of aging.
In 1993, dropping dietary methionine in rats from 0.86% to 0.17% extended median lifespan about 30% and maximum lifespan about 40%, with no early-mortality penalty. R
A second group replicated it with a 42% increase in mean lifespan and an 81 to 164% rise in blood glutathione. R
It works in mice too, where methionine restriction (MR) lowered serum IGF-1, insulin, glucose, and thyroxine while extending maximum lifespan. R
It does not even require starting young, since MR begun at 12 months of age (middle age in a mouse) still extended longevity. R
The mechanisms are where it gets useful.
MR raises Fibroblast Growth Factor 21 (FGF21), a fasting-state hormone, and FGF21 is required for the increase in energy expenditure and insulin sensitivity that MR produces (although it is not required for MR's effect on liver fat). R
MR drives the transsulfuration pathway to make more hydrogen sulfide (H2S), a gasotransmitter that is essential for dietary-restriction stress resistance across yeast, worms, flies, and rodents. R
MR downregulates mTORC1 through the SAM sensor described above, which shifts cells away from growth and toward maintenance. R
MR lowers mitochondrial reactive oxygen species generation and oxidative damage to mitochondrial DNA in rat liver and heart. R
MR even browns white fat through FGF21, increasing thermogenesis and energy burn. R
The metabolic payoff is real in the few human and applied-animal studies that exist.
In the only controlled human MR trial, 16 weeks of restriction in adults with metabolic syndrome shifted fuel use toward fat oxidation and reduced liver fat independent of weight loss. R
In obese mice, MR prevented and reversed hepatic steatosis by suppressing the de novo lipogenesis gene SCD1 and increasing fat export. R
There is a big MAYBE underneath all of this, though.
Almost every longevity number above is from rodents, and the single human trial measured metabolism over months, not lifespan over decades.
The H2S mechanism, as elegant as it is, is not unanimous, because at least one rat study found MR raised homocysteine and did not uniformly boost H2S-producing capacity. R
And MR in flies and yeast only extends lifespan when total amino acid intake is otherwise low, so it was never a standalone lever divorced from overall protein. R
That last caveat is the crack that the addition argument walks through.
The Case For Addition
The problem with "eat less methionine" as human advice is that the cleanest way to do it is to eat less protein, and eating less protein has its own bill.
Continuous MR in rodents reduces bone mineral density, limits lean mass, and stunts growth, which is exactly what you do not want in an aging human already losing muscle and bone. R
Cycling MR rather than running it continuously preserves more bone and lean mass and produces mechanically stronger bones, which tells you the continuous version is the harmful pattern. R
Methionine is also genuinely essential for the immune system, because T cells import methionine to make the SAM they need for the histone and RNA methylation that programs their activation. R
Strip methionine too far and you do not get a stress-resistant superorganism, you get a frail, sarcopenic, immunocompromised one.
This is the gap the newest research fills.
In June 2026, a team led by Maura Fanti and Valter Longo published a study in Cell Metabolism testing what happens when you take a low-protein, plant-and-fish longevity diet and add a small, controlled amount of methionine back in. R
They fed 20-month-old mice (roughly retirement age in human terms) one of four diets: standard chow, a Western high-fat high-sugar diet, a ketogenic diet, or the Longevity Diet with Methionine Modification (LDMM). R
The LDMM group lost fat without losing lean mass, reduced frailty, and improved cardiometabolic and insulin-sensitivity markers, and they did it while eating as many calories as any other group. R
The hormonal signature was the tell: LDMM raised growth hormone, raised Glucagon-Like Peptide 1 (GLP-1), raised FGF21, and lowered Insulin-like Growth Factor 1 (IGF-1). R
That is the same FGF21 and GLP-1 biology that makes the GLP-1 receptor drugs work, reached through food and a single amino acid instead of an injection.
Here is the line that matters most.
Too little methionine caused frailty, and too much methionine erased the diet's benefits entirely. R
The benefit lived in a narrow window the authors described as "low but sufficient," not at zero and not at maximum.
Longo's own summary was that amino acid composition, not just total protein quantity, is the lever, which is a direct challenge to the idea that you simply minimize protein or minimize methionine and call it longevity. R
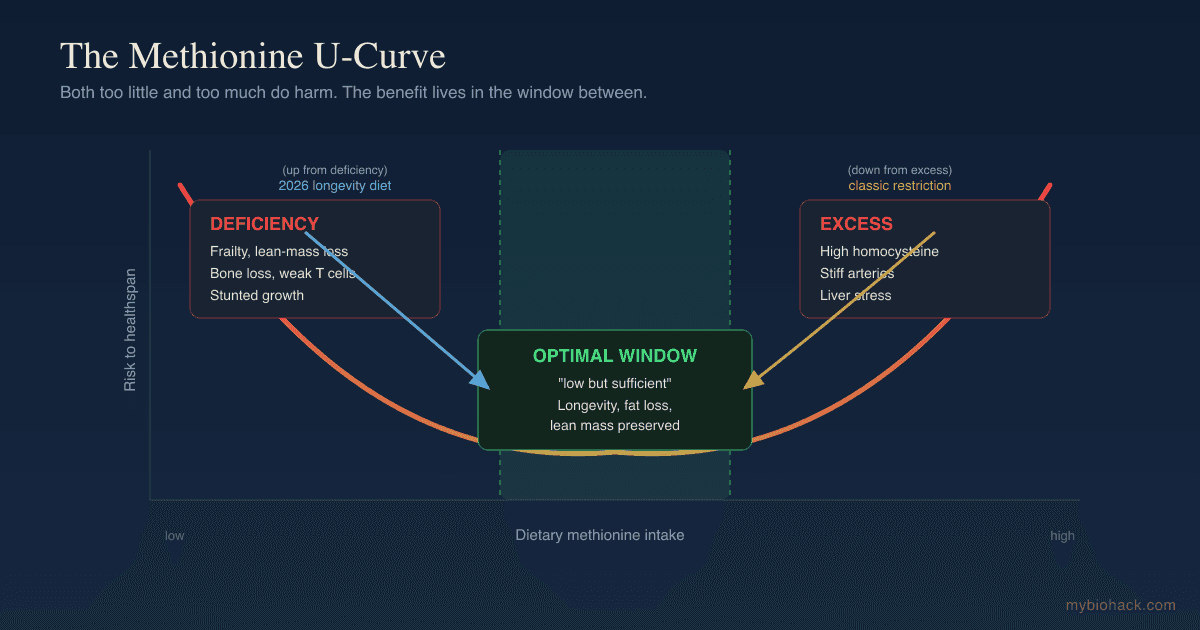
Reconciling The Two: The U-Shaped Curve
The restriction camp and the addition camp are not actually fighting, because they are describing two ends of the same curve.
Methionine has a U-shaped dose-response, where both deficiency and excess are harmful and the optimum is a window in between. R

At the low end you get frailty, lean-mass loss, bone loss, and impaired immunity.
At the high end you get the problems of excess, which the toxicity literature is blunt about: methionine is the most toxic amino acid when given well above need, largely because it floods the homocysteine pathway. R
A high-methionine diet in rats raised liver enzymes and homocysteine while depleting antioxidant defenses, a clear hepatotoxic signature. R
Most people reading this in a Western country are not methionine-deficient, because the average diet runs well above the requirement of roughly 19 mg/kg/day of total sulfur amino acids. R
The classic MR experiments slammed intake from the high shoulder of the curve down toward the trough, and benefit appeared because the starting point was excess.
The Fanti and Longo design started from the opposite problem, a low-protein diet already sitting near the deficient end, and benefit appeared by nudging methionine back up into the window. R
The reconciliation is dose, not direction.
This is the same shape you see with calorie restriction and fasting mimetics, where the goal is intermittent, dosed signaling rather than permanent maximal deprivation.
The Cancer Paradox
Methionine and cancer is where the "less is better" instinct gets genuinely dangerous if applied carelessly.
Most cancer cells are methionine-addicted, a phenomenon called the Hoffman effect, where they cannot grow on homocysteine alone even though normal cells can. R
Addicted cells burn methionine into excess and aberrant transmethylation, so they need a constant external supply to keep their histone methylation and proliferation running. R
This is why dietary methionine restriction looks like a cancer therapy in the right setting.
A landmark 2019 Nature study showed MR inhibited tumor growth and remodeled one-carbon metabolism in colorectal cancer xenografts and a sarcoma model, and it sensitized those tumors to low-dose chemotherapy and radiation. R
The same paper fed healthy humans a short methionine-restricted diet and showed it shifted the plasma one-carbon metabolome in the same direction, proving the diet engages the target pathway in people, not just mice. R
The pharmaceutical industry is chasing the same node from the other side, targeting MAT2A and PRMT5 in tumors with MTAP deletions. R
But there are two hard counterweights, and a responsible read keeps both in view.
First, methionine and folate can also fuel cancer rather than starve it, because a high-folate diet promoted liver cancer by stabilizing the methionine-cycle enzyme MAT IIα, and knocking that enzyme out abolished the effect. R
Second, and more important for anyone tempted to crash their methionine to fight a tumor, your T cells need methionine as badly as the tumor does.
Tumors physically outcompete CD8 T cells for methionine by overexpressing the transporter SLC43A2, and starving the T cells of methionine collapses their function and the response to immunotherapy. R
So whole-body methionine restriction can starve the tumor and disarm the immune system that is supposed to kill it, in the same stroke. R
This is not a do-it-yourself protocol.
Methionine restriction in active cancer is an adjunct studied alongside chemotherapy and radiation under supervision, not a substitute for treatment, and the human survival data do not yet exist.
If you are navigating this, it belongs in a conversation with your oncologist and is exactly the kind of nuance worth working through with a practitioner, which is what a consultation is for.
The Glycine Counterbalance
There is a way to blunt a high-methionine load without eating less protein, and it runs through glycine.
The liver disposes of excess SAM using glycine N-methyltransferase (GNMT), an enzyme so abundant it is one of the most common proteins in the liver, and its only job is to burn off surplus methyl groups by methylating glycine into sarcosine. R
Feeding rats excess methionine directly upregulates GNMT, which is the body demonstrating its own buffering system in real time. R
This is the basis of the McCarty and Brind hypothesis that glycine supplementation can act as a methionine-restriction mimetic, draining excess SAM through the GNMT overflow valve. R
The rodent longevity data are suggestive: glycine supplementation extended lifespan in genetically diverse mice in the NIA Interventions Testing Program. R
The practical translation is the methionine-to-glycine ratio of your diet.
Muscle meat is high in methionine and low in glycine, while the glycine sits in the parts most people throw away: skin, connective tissue, and bone.
Eating nose-to-tail, or adding glycine or collagen (which is roughly one-third glycine), restores a ratio that muscle-meat-heavy eating distorts.
The honest caveat is that the lifespan-mimicry evidence is rodent-only, and no human trial has shown glycine reproduces the benefits of methionine restriction.
It is a low-risk, biologically grounded hedge, not a proven longevity drug.
Who Should Restrict And Who Should Add
The U-shape means the right move depends entirely on where you already sit, so this is not one recommendation, it is two.
Methionine reduction (or glycine balancing) makes more sense if you are: (not an exhaustive list)
- Carrying metabolic syndrome, fatty liver, or excess fat mass, where short methionine and cysteine restriction raised FGF21 and improved metabolic markers in a human pilot R
- Eating a muscle-meat-heavy diet with little connective tissue, glycine, or collagen
- Optimizing for longevity from a starting point of high protein intake
- Working with an oncologist on an adjunct protocol for a methionine-dependent cancer R
Methionine sufficiency (eating enough, not restricting) makes more sense if you are: (not an exhaustive list)
- Eating a vegan or low-protein diet, which is inherently the lowest in methionine and sulfur amino acids R
- Fighting an active infection or relying on a functioning T-cell compartment R
- Frail, older, or losing muscle, where too-little methionine drove frailty in the Fanti and Longo model and where preserving lean mass matters more than squeezing out a longevity signal R
- Recovering from surgery, illness, or anything demanding active tissue building
The overlap case is the most common one in real life: a middle-aged person who is metabolically a little off but does not want to lose muscle.
For that person the answer is rarely "restrict methionine" and almost always "fix the protein source, add the glycine, and dose the signaling with fasting rather than chronic deprivation."
What To Stay Away From
Do not megadose L-methionine as a standalone supplement chasing a vague "methylation" or "detox" benefit.
Oral methionine supplementation has thin benefit evidence and reliably raises homocysteine, which is the opposite of what you want. R
An oral methionine load acutely impairs endothelium-dependent vasodilation in healthy adults, the same flow-mediated dilation that depends on nitric oxide. R
It also acutely stiffens arteries, dropping systemic arterial compliance by roughly 22% within hours. R
Do not run continuous, aggressive methionine restriction long-term without watching bone and lean mass, because the continuous pattern is the one that costs you skeleton and muscle. R
Do not attempt methionine restriction as a cancer treatment on your own, for the T-cell reason above. R
Do not restrict methionine hard if you are also low in B12, folate, or B6, because you will strain the same remethylation and transsulfuration machinery that keeps homocysteine in check. R
Testing
You cannot dose methionine intelligently without knowing where you sit on the cycle, and most of that is a blood draw.
Blood And Urine Markers
Homocysteine is the single most useful readout, because it integrates your methionine load against your B12, folate, and B6 status, and elevated homocysteine is associated with higher cardiovascular and stroke risk in large meta-analyses (optimal is generally under 7 to 8 µmol/L, with risk rising above 10). R
I use the Homocysteine + B12 + Folate panel (Quest) to read homocysteine against the cofactors that actually control it, rather than testing it in isolation.
For the cofactors individually, the Vitamin B12 and Vitamin B6 tests (Quest) round out the picture, since B6 specifically gates the transsulfuration exit.
Functional Lab Panels
I use the Nutrient Zoomer (Vibrant Wellness) to assess B6, B12, folate, zinc, and the amino acid panel including methionine and cysteine in one pass.
I use the Cellular Zoomer (Vibrant Wellness) to read organic acids and methylation markers, which show how the one-carbon cycle is actually running rather than just what is in the tank.
For metabolic context, since the entire addition argument hinges on insulin sensitivity and fat mass, I use the Cardio Zoomer (Vibrant Wellness) to assess fasting insulin, ApoB, and the lipid and metabolic markers that move with methionine status.
Genetics
I use the Methylation Genetics panel (Vibrant Wellness) to read MTHFR, MTR, MTRR, BHMT, and CBS together, because your variants change how much methionine load your remethylation and transsulfuration pathways can absorb.
If you only want the single highest-yield variant, the MTHFR DNA Mutation Analysis (Quest) covers C677T and A1298C on their own.
Mechanisms Of Action
Simple:
- Methionine becomes SAM, the molecule your cells use both to methylate everything and to sense whether there is enough food around to grow.
- When methionine is high your growth pathway switches on, and when it is low your repair and stress-resistance pathways switch on instead.
- Too little leaves you frail and immune-compromised, too much floods you with homocysteine and stiffens your arteries, and the benefit lives in the window between.
Advanced:
- SAM as the mTORC1 signal. Methionine is sensed indirectly through SAM, where SAMTOR binds SAM and, as SAM falls, releases to activate GATOR1 and shut down mTORC1; SAM, not methionine itself, is the molecular signal, which is why anything that drains SAM (low methionine, high GNMT flux from glycine) mimics restriction. R
- FGF21 induction. Methionine restriction raises hepatic FGF21, which is required for the increase in energy expenditure, white-adipose browning, and insulin sensitization, but is dispensable for the hepatic lipid effect, so MR's benefits are not routed through one master switch. R
- Transsulfuration and H2S. Sulfur-amino-acid restriction increases cystathionine gamma-lyase flux and endogenous hydrogen sulfide, which is required for dietary-restriction stress resistance, while CBS is allosterically activated by SAM so high methylation status itself pushes homocysteine toward this branch. R R
- SCD1 suppression. A core driver of MR's triglyceride-lowering and anti-steatosis effect is suppression of stearoyl-CoA desaturase-1, a de novo lipogenesis gene, which redirects the liver from making fat to exporting and oxidizing it. R
- GNMT as the SAM rheostat. Glycine N-methyltransferase burns surplus SAM by methylating glycine to sarcosine with no useful product, setting the ceiling on SAM, which is the mechanistic basis for glycine acting as a restriction mimetic and for the methionine-to-glycine dietary ratio mattering. R
Genetics
MTHFR
MTHFR encodes the enzyme that produces 5-methyltetrahydrofolate, the folate form that MTR uses to remethylate homocysteine back to methionine.
Variants that slow it raise homocysteine, especially when folate or riboflavin status is low.
rs1801133 (C677T, Ala222Val) — the thermolabile variant, where the TT genotype reduces enzyme activity by roughly 70% and raises plasma homocysteine. R
rs1801131 (A1298C, Glu429Ala) — a milder variant with a weaker and less consistent effect on homocysteine, though compound heterozygosity with C677T can lower activity more than either alone. R
Targeted riboflavin lowered blood pressure specifically in 677TT individuals, a genotype-specific effect not seen in other genotypes, because the variant binds its FAD cofactor less tightly. R
This variant also sits at the center of the histamine and methylation interplay covered in naturally treating histamine intolerance and the A1298C and BH4 discussion.
MTR / MTRR
MTR encodes methionine synthase, the B12-dependent enzyme that remethylates homocysteine, and MTRR keeps it active by regenerating its cobalamin cofactor.
Variants modify remethylation efficiency and homocysteine, with effects smaller and more context-dependent than MTHFR C677T.
rs1805087 (MTR A2756G) and rs1801394 (MTRR A66G) — modest modifiers of the remethylation arm that matter most when B12 is marginal. R
CBS
CBS encodes cystathionine beta-synthase, the B6-dependent gate that commits homocysteine irreversibly to transsulfuration toward cysteine, glutathione, and H2S.
Loss-of-function mutations cause classical homocystinuria, with very high homocysteine, treated with B6, a low-methionine diet, and betaine.
CBS is allosterically activated by SAM, which is the elegant part: a high methylation load itself opens the transsulfuration exit, so the cycle has a built-in pressure valve. R
GNMT
GNMT encodes glycine N-methyltransferase, the liver's dedicated SAM overflow valve.
Loss-of-function mutations cause persistent hypermethioninemia with mild liver disease, and the knockout mouse accumulates extreme SAM, confirming GNMT sets the SAM ceiling.
Characterized human mutations include L49P (about 10% residual activity) and N140S (under 0.5%). R
MAT1A / MAT2A
MAT1A encodes the liver-specific enzyme that makes SAM, while MAT2A is the version expressed in all other tissues and in proliferating or dedifferentiated cells.
The switch from MAT1A to MAT2A is a hallmark of liver cancer and is the node the new MAT2A inhibitor drugs target. R
MAT1A deficiency causes usually-benign isolated hypermethioninemia, while MAT2A upregulation marks the proliferative state. R
More Research
For biomarker testing I use the Homocysteine + B12 + Folate panel and the Cellular Zoomer to track the one-carbon cycle over time inside the Health Hub, which is the right place to watch a marker like homocysteine respond to a dietary change.
Intermittent rather than continuous methionine restriction preserves bone and lean mass while keeping the metabolic benefit, which mirrors the broader principle that pulsed signaling beats chronic deprivation. R
Methionine restriction's reproductive cost is separable from its longevity benefit, since folic acid rescued fertility in flies while preserving the lifespan extension, which means the two effects run on different tracks. R
NAC, or N-acetylcysteine, supplies cysteine and spares the methionine requirement, which is one way to support glutathione without leaning on the methionine cycle. R
SAMe supplementation has the best evidence as an adjunct for depression at 1,600 to 3,200 mg/day and for SAM-depleted liver states, though monotherapy and osteoarthritis data are mixed. R
The acute methionine-load vascular effect is not universal, since at least one controlled study found induced homocysteine changes did not impair flow-mediated dilation in healthy volunteers, so the endothelial story is strong but not unanimous. R
The whole field is U-shaped, which is the one durable takeaway: both deficiency and excess are harmful, "optimal" methionine is a window rather than a number to maximize or minimize, and the human longevity data for either direction do not yet exist. R
Jacob Gordon
INHC, FMT-C
Board Certified Health Coach
I spent years battling unexplained chronic illness before discovering biohacking, epigenetics, and functional medicine. Now I share that research at MyBioHack to help others find their own answers.
Book a ConsultationRelated Protocols & Supplements
Deep-dive chapters and recommended supplements for this topic
Quercetin
500mg 2x/day
Vitamin D3 + K2
5000 IU + 200mcg/day
Magnesium Glycinate
400mg at bedtime