The Epigenetics Of Attachment Style: A Gene-By-Gene Map
By Jacob Gordon, INHC, FMT-CThis article contains affiliate links. As an Amazon Associate, MyBioHack earns from qualifying purchases at no extra cost to you. We only link products we research and stand behind.
Attachment style is the set of patterns you run in close relationships, and the internet increasingly sells it as a fixed readout of your genes, which is mostly wrong but not entirely empty.
In this post, we will discuss every major gene and pathway people invoke when they talk about the biology of attachment, build an honest case for and against each one, and separate the handful with real evidence from the larger group that is inferred, indirect, or already broken by the candidate-gene replication crisis.

How To Read This Map
The honest starting point is that attachment style is a learned relational pattern, not a genotype.
It is built in the first years of life through coregulation with caregivers, and the genes below modulate how that learning lands, not whether it happens.
So treat this map as a list of dials that bias the system, not switches that set your relationships.
There are three recurring problems with almost everything that follows, and naming them once saves repeating them under every gene.
The first is the candidate-gene replication crisis.
When researchers tested the most famous psychiatric candidate genes across samples of tens to hundreds of thousands of people, none of the historical single-gene hypotheses held up, and the effects that launched a thousand blog posts mostly evaporated. R
The second is that gene-by-environment interaction studies, which is where most attachment genetics lives, replicated at a dismal rate, with 96% of novel findings significant versus 27% of replication attempts, a signature of underpowered studies and false positives. R
The third is polygenicity, meaning any trait this complex is shaped by thousands of variants each with a vanishingly small effect, so a single SNP almost never carries the weight people assign it. R
The most defensible frame for the whole field is differential susceptibility, where a so-called risk allele is better understood as a plasticity allele that makes you more shaped by your environment for better and for worse. R
The same genotype that does worst under harsh caregiving often does best under warm caregiving, which means these variants index sensitivity to relationships, not a fixed relational fate. R
Where a gene's attachment link runs through epigenetics rather than the DNA sequence, that is usually the stronger story, because early caregiving writes methylation marks onto these genes that persist into adulthood.
Read each entry as "here is the strongest case anyone can make, and here is why it is weaker than it sounds."
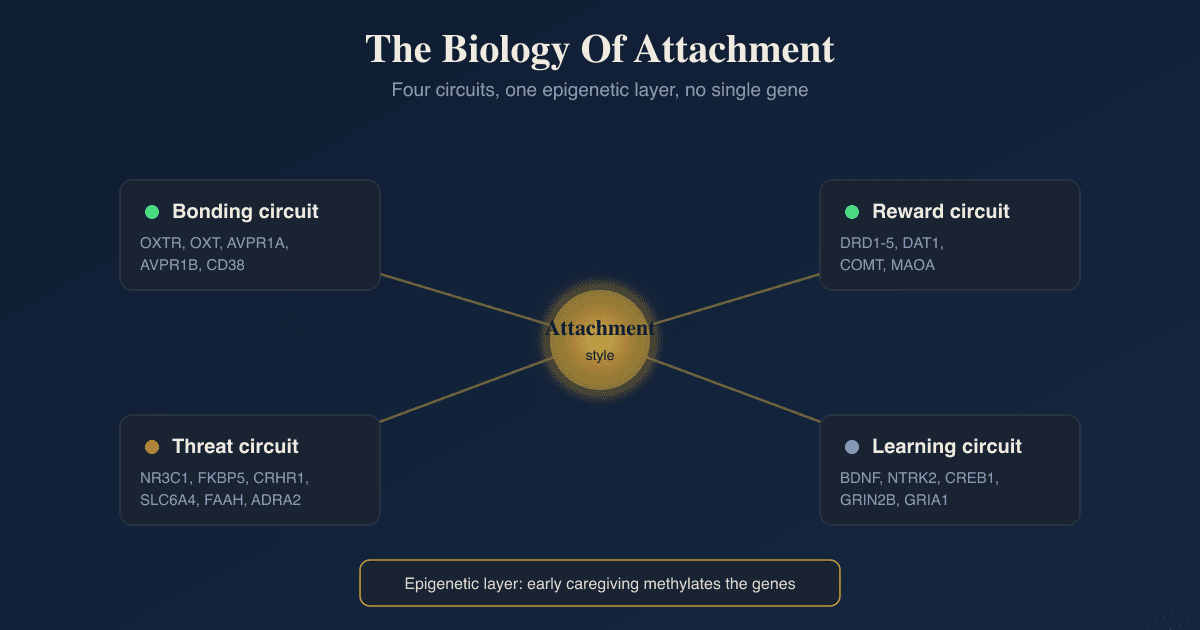
Social Bonding Systems
This is the cluster with the most direct claim to attachment, because oxytocin and vasopressin are the neuropeptides that physically tag closeness, trust, and pair bonding.
It is also the cluster where the epigenetic evidence is genuinely better than the SNP evidence.
OXTR, the Oxytocin Receptor (rs53576, rs2254298, ligand oxytocin).
For: OXTR is the docking site for the main bonding signal, its promoter methylation tracks attachment directly, where higher methylation associates with more attachment avoidance and lower methylation with less attachment anxiety and higher plasma oxytocin. R R
For: that methylation is written early, since childhood maltreatment and low maternal care both raise OXTR methylation in ways that last, which is exactly the mechanism you would want for "early relationships shape adult attachment."
Against: the famous rs53576 SNP itself is the weak part, since the largest meta-analysis covering 48 effect sizes in over 17,000 people found combined effects on sociality that did not differ from zero. R
Against: a separate meta-analysis salvaged a small general-sociality signal for rs53576 but found it unrelated to close relationships specifically, which is the opposite of what the attachment framing needs. R
The verdict is that OXTR methylation is one of the cleanest molecular handles on attachment in this entire map, while the rs53576 genotype is mostly noise.
OXT, the Oxytocin gene (rs2740210, ligand oxytocin).
For: OXT encodes oxytocin itself, OXT methylation has been linked to lower plasma oxytocin and to less secure attachment, and rs2740210 sits in a regulatory region tied to social and parenting behavior.
Against: the human OXT SNP literature is small, inconsistent, and dominated by candidate-gene studies of the kind that rarely replicate, so the production gene is a weaker handle than the receptor gene.
The verdict is plausible and mechanistically central, but thin on its own.
AVPR1A, the Vasopressin Receptor 1A (RS3 repeat, rs11174811, ligand vasopressin).
For: this is the human echo of the vole pair-bonding story, and the RS3 repeat length has been associated with partner bonding, perceived marital problems, and marital status in men, with a dose-dependent effect of the 334 allele. R
For: AVPR1A variation has also been tied to trust and reciprocity in economic-game paradigms. R
Against: the original pair-bonding effect was male-specific, modest, and has had an inconsistent replication record, and the leap from vole microsatellites to human marriage is exactly the kind of just-so story the field has learned to distrust.
The verdict is one of the better-known bonding findings, real but small and shaky.
AVPR1B, the Vasopressin Receptor 1B (rs28632197, ligand vasopressin).
For: AVPR1B sits at the pituitary end of the HPA axis and shapes how social stress and aggression are processed, which is upstream of how safe closeness feels.
Against: there is essentially no direct human attachment-style data, the link is inferred from stress and aggression phenotypes, and it inherits all the candidate-gene replication problems. R
The verdict is mechanistically reasonable, empirically absent for attachment.
CD38 (rs3796863, regulates oxytocin release).
For: CD38 is genuinely underrated, because it controls oxytocin secretion from neurons, and knocking it out in mice cripples social behavior in a way that oxytocin rescues. R
For: in humans, the rs3796863 risk allele shifts neural processing of social stimuli and interacts with intranasal oxytocin, plus it has been associated with autism in two family samples. R
Against: the human work is small and centered on autism and lab social-processing tasks rather than attachment style as such, so the attachment claim is an extrapolation.
The verdict is a strong mechanism for oxytocin availability, with attachment inferred rather than measured.
Dopamine And Reward
Bonding is not only a peptide event, it is a reward event, and dopamine is what makes closeness feel worth seeking and repeating.
This is why the dopamine genes are the second-strongest cluster, and also where differential susceptibility was first demonstrated genetically.
DRD1 (rs4532, dopamine).
For: D1 receptors drive reward learning and have a role in the formation of partner preference in animal bonding models, so the gene is mechanistically in the circuit.
Against: human DRD1 attachment data is sparse to nonexistent, and rs4532 is a generic candidate SNP with the usual replication weakness.
The verdict is circuit-relevant, attachment-unproven.
DRD2 (rs1800497, dopamine).
For: the D2 receptor is required, together with oxytocin, for pair-bond formation in the nucleus accumbens, making it one of the most mechanistically justified genes here. R
For: the rs1800497 Taq1A allele lowers striatal D2 density and is repeatedly tied to reward sensitivity and addiction, which plausibly colors how rewarding closeness is.
Against: rs1800497 is not even in DRD2, it sits in the neighboring ANKK1 gene, and its attachment relevance is inferred from reward phenotypes rather than measured against attachment style.
The verdict is strong mechanism, indirect human evidence.
DRD3 (rs6280, dopamine).
For: D3 receptors concentrate in limbic reward regions and shape emotional salience, the weighting of what feels important about a social partner.
Against: no meaningful human attachment literature, pure inference from receptor distribution.
The verdict is speculative.
DRD4 (7-repeat VNTR, dopamine).
For: this is the historical home of genetic differential susceptibility, where the 7-repeat allele made children more shaped by parenting for better and for worse, more disorganized attachment under insensitive care and more benefit from sensitive care. R
For: the meta-analysis of dopamine-related genes supports the plasticity reading over a simple risk reading. R
Against: several of the original DRD4 differential-susceptibility findings have failed to replicate in larger samples, and the effect, if real, is small and environment-dependent rather than a standalone attachment gene.
The verdict is the most theoretically important dopamine variant here, with the plasticity frame more durable than any single result.
DRD5 (rs6283, dopamine).
For: D5 supports reward signaling and is part of the same circuit.
Against: negligible human attachment evidence, candidate-SNP weakness.
The verdict is filler unless paired with the broader dopamine picture.
DAT1 / SLC6A3 (40bp VNTR, dopamine clearance).
For: the dopamine transporter sets how long dopamine lingers in striatal reward circuits, so it tunes the gain on social reward, and it has appeared in differential-susceptibility models alongside DRD4. R
Against: the attachment link is entirely through reward and ADHD phenotypes, not attachment style directly, and replication is inconsistent.
The verdict is a reasonable tuning knob, indirectly evidenced.
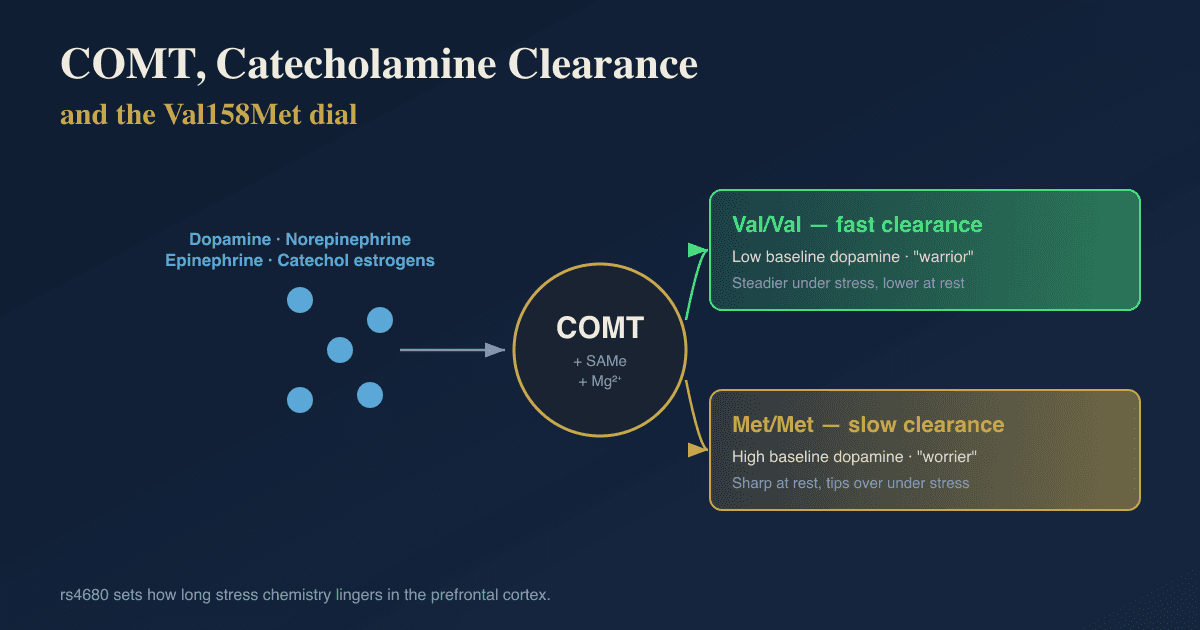
COMT (rs4680 Val158Met, dopamine and norepinephrine).
For: COMT clears prefrontal dopamine and norepinephrine, the Met allele has a direct published link to avoidant attachment, and its promoter methylation tracks impaired fear inhibition that overlaps with insecure attachment, covered fully in the COMT and attachment post.
Against: the attachment effect followed a positive-heterosis pattern where heterozygotes scored highest, not the tidy "Met is anxious" story, and the broader COMT-personality literature has small, sex-dependent, often-flipping effects.
The verdict is one of the few catecholamine genes with a direct attachment paper, but the effect is non-linear and modest.
MAOA (uVNTR, dopamine, serotonin, norepinephrine).
For: MAOA degrades all three monoamines, and the low-activity allele combined with childhood maltreatment is one of the most cited gene-by-environment findings in psychiatry, predicting later antisocial and aggressive outcomes that bleed into relational security.
Against: the original Caspi MAOA finding has a mixed replication record, and it is a violence-and-aggression story retrofitted onto attachment rather than an attachment finding, with all the G×E fragility named earlier. R
The verdict is famous, mechanistically broad, and shakier than its fame suggests.
Serotonin
Serotonin tunes rejection sensitivity, threat appraisal, and the affective tone you bring to social signals.
It is also the single best cautionary tale in this entire field.
SLC6A4, the serotonin transporter (5-HTTLPR, rs25531, serotonin).
For: the short allele of 5-HTTLPR was the original gene-by-environment superstar, where carriers showed more depression after stressful life events, and it has been tied to rejection sensitivity and amygdala threat reactivity. R
Against: the landmark replication attempt, a meta-analysis of 14 studies, found no evidence that 5-HTTLPR alone or with stressful life events raised depression risk, and this became the textbook example of a candidate-gene finding that did not hold. R
The verdict is that 5-HTTLPR is the most overstated variant in psychology, useful mostly as a warning about how confident the early literature sounded.
HTR1A (rs6295, serotonin).
For: the 5-HT1A autoreceptor sets serotonergic tone and anxiety regulation, and the G allele has been linked to anxiety and depression phenotypes that shape relational reactivity.
Against: no direct attachment data, and the anxiety associations carry the standard candidate-gene caveats. R
The verdict is plausible for the anxious end of the spectrum, unproven for attachment.
HTR1B (rs6296, serotonin).
For: 5-HT1B shapes aggression and social behavior in animal models.
Against: essentially no human attachment-style evidence.
The verdict is speculative.
HTR2A (rs6311, rs6313, serotonin).
For: 5-HT2A is central to emotional processing and was a heavily studied depression candidate.
Against: it was one of the genes specifically debunked in the large candidate-gene re-examination, so the case against is unusually strong. R
The verdict is mechanistically interesting, empirically discredited as a main effect.
HTR2C (rs6318, serotonin).
For: 5-HT2C influences stress sensitivity and mood, and the gene is X-linked, which makes for interesting sex differences.
Against: no attachment-specific evidence, candidate-SNP fragility.
The verdict is weak.
TPH2 (rs4570625, serotonin synthesis).
For: TPH2 is the brain's rate-limiting enzyme for serotonin synthesis, so variants plausibly set baseline serotonergic capacity and emotional regulation.
Against: human behavioral associations are inconsistent and there is no clean attachment link.
The verdict is upstream and important biologically, unproven behaviorally.
Cortisol And The HPA Axis
This is the cluster where epigenetics, not the SNP, carries the strongest and most replicated attachment-relevant signal.
The reason is simple, because attachment is fundamentally about whether other people regulate your stress system, which is exactly what the HPA axis does, and the same chronically activated stress system shows up across insecure attachment.
NR3C1, the glucocorticoid receptor (rs6198, rs41423247, cortisol).
For: this is the flagship attachment-epigenetics finding, where childhood abuse was associated with increased methylation of the NR3C1 promoter in human brain, lowering receptor expression and blunting the off-switch on the stress response, a direct molecular trace of early adversity. R
For: NR3C1 promoter methylation has since been specifically associated with attachment avoidance in young adults, linking the mark to attachment style and not just to abuse. R
Against: some NR3C1 methylation findings have been hard to replicate and effect sizes are small, and the SNPs themselves are far weaker than the methylation story.
The verdict is the single most important entry in this map, with the epigenetic evidence strong and the genetic evidence secondary.
NR3C2, the mineralocorticoid receptor (rs5522, cortisol).
For: the MR sets baseline tonic stress regulation and the appraisal of threat, and the rs5522 variant has been tied to dispositional optimism and stress reactivity.
Against: no direct attachment-style data, modest and inconsistent behavioral findings.
The verdict is mechanistically sensible, empirically thin for attachment.
FKBP5 (rs1360780, rs3800373, cortisol).
For: FKBP5 is a strong gene-by-environment story, where the rs1360780 T allele combined with childhood abuse raised PTSD risk and altered HPA-axis feedback, and the interaction is partly mediated by demethylation of FKBP5 after early trauma. R
For: that trauma-driven epigenetic change to a stress gene is precisely the kind of mechanism that could lock in a threat-biased relational style.
Against: like all G×E findings the replication is imperfect, and the phenotype studied is PTSD and stress reactivity rather than attachment style directly. R
The verdict is one of the better-supported stress-gene-by-trauma interactions, attachment-adjacent rather than attachment-specific.
CRH (rs28364015, cortisol pathway).
For: CRH initiates the entire stress cascade and is the upstream signal that caregiving either amplifies or dampens.
Against: human variant data tied to attachment is essentially absent.
The verdict is upstream and central, empirically quiet.
CRHR1 (rs110402, cortisol pathway).
For: CRHR1 has one of the more striking findings in the field, where protective haplotypes including the rs110402 A allele buffered against adult depression following childhood maltreatment, a real gene-by-environment effect with at least one replication. R
Against: the protective effect was sex-specific, appearing in men but not women in some samples, which limits how cleanly it generalizes. R
The verdict is a credible buffering variant for the trauma-to-depression path, with attachment inferred.
CRHR2 (rs2267717, cortisol pathway).
For: CRHR2 is involved in stress recovery and social-stress buffering, the recovery half of the threat response.
Against: minimal human attachment data.
The verdict is plausible, unproven.
POMC (rs1866146, ACTH and beta-endorphin).
For: POMC sits at the crossroads of the cortisol axis and the endorphin system, so it touches both stress regulation and the opioid comfort of closeness.
Against: no attachment-specific human evidence.
The verdict is mechanistically interesting for bridging two systems, empirically absent.
Opioids And Social Pain
The opioid system is the biology of social comfort and the sting of rejection, which is conceptually closer to attachment than almost anything else here.
This is the brain-opioid theory of social attachment, that the same system handling physical pain handles the pain of separation.
OPRM1, the mu-opioid receptor (rs1799971 A118G, endorphins).
For: the A118G variant was associated with dispositional and neural sensitivity to social rejection, where G carriers reported more rejection sensitivity and showed greater activity in social-pain regions. R
For: the same variant moderated how early maternal care translated into fearful attachment, a direct bridge from caregiving to an attachment trait. R
Against: a larger preregistered replication with 490 people failed to find that OPRM1 moderates social-rejection sensitivity, which seriously undercuts the original. R
The verdict is conceptually the best-fitting gene in the map and one of the better-replicated cautionary failures, a perfect illustration of how compelling and how fragile this field is at once.
OPRD1, the delta-opioid receptor (rs2234918, endorphins).
For: delta-opioid signaling supports emotional resilience and mood stability, part of the buffering tone closeness provides.
Against: no human attachment-style data.
The verdict is speculative.
OPRK1, the kappa-opioid receptor (rs1051660, dynorphin).
For: the kappa-dynorphin system drives dysphoria and the aversive side of stress, and is mechanistically tied to the distress of separation and social loss.
Against: human attachment evidence is absent, the link is from animal dysphoria models.
The verdict is mechanistically apt for attachment distress, unmeasured in people.
PENK, proenkephalin (rs2609997, enkephalins).
For: enkephalins mediate social soothing and comfort, and PENK expression responds to social contact in animal models.
Against: no human attachment-style genetics.
The verdict is plausible, unproven.
Norepinephrine And Vigilance
Norepinephrine sets vigilance, and hypervigilance is a core feature of both anxious and disorganized attachment.
The case here is mostly mechanistic, since these genes are studied for arousal and PTSD rather than attachment.
ADRA2A (rs1800544, norepinephrine).
For: the alpha-2A autoreceptor governs noradrenergic tone and vigilance, plausibly tuning the hyperarousal of insecure attachment.
Against: no direct attachment data, candidate-SNP weakness.
The verdict is mechanistically on point, empirically thin.
ADRA2B (deletion variant, norepinephrine).
For: the ADRA2B deletion is one of the cleaner human findings here, robustly enhancing emotional memory so that emotionally charged events, including relational ones, are encoded more vividly.
Against: enhanced emotional memory is a general affective trait, not attachment, so the attachment relevance is an extrapolation.
The verdict is a real cognitive effect, indirectly relevant.
ADRB1 (rs1801253, norepinephrine).
For: beta-1 adrenergic signaling shapes stress arousal and cardiovascular reactivity to social stress.
Against: no attachment-specific evidence.
The verdict is weak.
ADRB2 (rs1042713, rs1042714, norepinephrine).
For: beta-2 adrenergic variants tune fight-or-flight intensity and have been studied in anxiety and stress reactivity.
Against: no attachment link, generic candidate SNPs.
The verdict is weak.
The Endocannabinoid System
The endocannabinoid system buffers anxiety, supports social reward, and governs how flexibly you recover from stress, all covered in depth in the endocannabinoid system post.
It is an underappreciated emotional-regulation layer, with one genuinely strong human variant.
CNR1, the CB1 receptor (rs1049353, anandamide).
For: CB1 mediates social reward and emotional flexibility, and CNR1 variants have been tied to social reward responsiveness and to how strongly faces are processed as rewarding.
Against: human findings are small and not attachment-specific.
The verdict is mechanistically relevant, modestly evidenced.
CNR2, the CB2 receptor (rs2501432, endocannabinoids).
For: CB2 sits at the immune-neural interface and could link inflammation to social behavior.
Against: almost no human behavioral data, let alone attachment.
The verdict is speculative.
FAAH (rs324420 C385A, anandamide).
For: this is the standout, because the low-activity 385A allele raises anandamide, and carriers show quicker amygdala habituation to threat, stronger fronto-amygdala connectivity, faster fear extinction, and lower stress-reactivity, exactly the threat-processing profile that underlies felt safety in relationships. R R
Against: it is a threat-and-extinction finding rather than a measured attachment effect, so the attachment link is mechanistic inference, strong as that inference is.
The verdict is one of the most biologically convincing entries here for the felt-safety dimension, even without a direct attachment study.
MGLL, monoacylglycerol lipase (rs604300, 2-AG).
For: MGLL controls 2-AG tone and stress recovery, and rs604300 has interacted with childhood adversity in addiction phenotypes.
Against: no attachment data, the evidence is in stress and substance use.
The verdict is plausible, indirect.
GABA
GABA is the brain's primary brake, and the capacity to self-soothe and inhibit a threat response is close to the physiological core of secure attachment.
The genes are studied for anxiety and alcohol, not attachment, so this whole cluster is mechanistic inference.
GABRA2 (rs279858, GABA).
For: GABRA2 is well studied in anxiety and alcohol dependence, plausibly setting baseline anxiety tone and the ability to calm down after activation.
Against: no attachment-style data, and the alcohol-dependence associations have their own replication debates.
The verdict is mechanistically reasonable, empirically unproven for attachment.
GABRB3 (rs1426217, GABA).
For: GABRB3 has been studied in autism and emotional inhibition, which touches social regulation.
Against: no attachment-specific evidence.
The verdict is weak.
GABRG2 (rs211037, GABA).
For: the gamma-2 subunit is required for synaptic GABA-A function and nervous-system calming.
Against: studied mainly in epilepsy and anxiety, with no attachment link.
The verdict is weak.
Glutamate
Glutamate drives emotional learning and memory, including the learning of who is safe, which is arguably the substrate attachment is built on.
This cluster is probably underexplored, but that means the evidence is thin rather than negative.
GRIN2B (rs1806201, glutamate).
For: the NMDA receptor subunit GluN2B is central to synaptic plasticity and emotional learning, the machinery that encodes attachment experiences.
Against: no human attachment-style genetics, pure mechanistic inference.
The verdict is theoretically important, empirically blank.
GRM3 (rs6465084, glutamate).
For: the metabotropic receptor mGluR3 is involved in social cognition and has been studied in schizophrenia.
Against: no attachment evidence.
The verdict is speculative.
GRIA1 (rs548294, glutamate).
For: AMPA-receptor signaling underlies the consolidation of emotional memory, including relational memory.
Against: no attachment-specific data.
The verdict is mechanistically plausible, unstudied for attachment.
Neuroplasticity
Attachment is learned, so the genes governing how readily the brain rewires are upstream of whether attachment patterns can form and change, including in therapy, supported by the same biology as the BDNF post.
BDNF (rs6265 Val66Met, BDNF).
For: the Met allele reduces activity-dependent BDNF release, blunting plasticity, and it interacts with childhood trauma to alter limbic gray matter, a plausible route by which early adversity gets structurally embedded. R
For: lower plasticity could make attachment patterns, once set, harder to update.
Against: BDNF Val66Met was another gene flagged in the large candidate-gene re-examination as not holding up as a main effect for mood, so its standalone effect is small. R
The verdict is a real plasticity variant with a credible trauma interaction, weak as a direct attachment gene.
NTRK2, the BDNF receptor TrkB (rs1187323, BDNF receptor).
For: TrkB transduces the BDNF signal, so it is mechanistically inseparable from plasticity and emotional adaptation.
Against: no direct attachment data.
The verdict is mechanistically necessary, empirically quiet.
CREB1 (rs2253206, CREB).
For: CREB consolidates long-term memory, including the memory of attachment experiences, and is downstream of most plasticity signaling.
Against: no attachment-specific human genetics.
The verdict is foundational biology, unmeasured for attachment.
Sex Hormones
Sex hormones modulate oxytocin sensitivity and social behavior, which is part of why so many findings in this map differ by sex, and they interact with estrogen metabolism.
ESR1, estrogen receptor alpha (rs2234693, rs9340799, estrogen).
For: estrogen upregulates oxytocin receptor expression, so ESR1 variation plausibly sets oxytocin sensitivity, and ESR1 has been linked to anxiety and to OXTR expression.
Against: the attachment link is two steps removed, through estrogen then oxytocin, with no direct attachment study.
The verdict is a credible modulator of the bonding system, indirectly evidenced.
ESR2, estrogen receptor beta (rs1256049, estrogen).
For: ER-beta is involved in anxiety and social behavior in animal models.
Against: no human attachment data.
The verdict is speculative.
AR, the androgen receptor (CAG repeat, testosterone).
For: shorter CAG repeats mean a more sensitive androgen receptor, and androgen tone shifts dominance, bonding, and the trade-off between mating and parenting effort.
Against: the attachment relevance is inferred from dominance and pair-bonding trade-offs, not measured as attachment style.
The verdict is mechanistically interesting for the avoidant-dismissing end, unproven.
PGR, the progesterone receptor (rs1042838, progesterone).
For: progesterone supports nurturing and affiliative behavior, especially around pregnancy.
Against: no attachment-style genetics.
The verdict is weak.
CYP19A1, aromatase (rs700518, estrogen synthesis).
For: aromatase converts testosterone to estrogen and so sets the local estrogen that modulates bonding circuits.
Against: no direct attachment evidence.
The verdict is an upstream modulator, empirically blank.
The Cholinergic System
Acetylcholine governs social attention, the orienting toward and reading of social cues that attachment depends on.
This cluster is studied for attention and autism, not attachment.
CHRNA4 (rs1044396, acetylcholine).
For: the alpha-4 nicotinic subunit shapes attention, and social attunement starts with attending to the right social signals.
Against: no attachment-specific data.
The verdict is mechanistically tangential, unproven.
CHRNA7 (rs2337506, acetylcholine).
For: the alpha-7 subunit is studied in schizophrenia and autism for sensory and social information processing.
Against: no attachment-style evidence.
The verdict is weak.
Inflammation
Inflammation and social behavior run in a loop, where loneliness and social threat raise inflammatory tone and inflammation in turn promotes social withdrawal, part of Jacob's broader systemic inflammation picture.
The genes are studied for inflammation, with social behavior as a downstream correlate.
IL6 (rs1800795, IL-6).
For: IL-6 rises with social threat and loneliness, and the rs1800795 promoter variant sets IL-6 production, plausibly tuning social-threat sensitivity.
Against: this is a social-stress-physiology link, not an attachment-style finding, and the SNP-to-behavior chain is long.
The verdict is mechanistically real for the social-threat loop, indirect for attachment.
TNF (rs1800629, TNF-alpha).
For: TNF-alpha is induced by social stress and contributes to the sickness-behavior pattern of withdrawal.
Against: no attachment-specific data.
The verdict is indirect.
IL1B (rs16944, IL-1 beta).
For: IL-1 beta drives social withdrawal in the sickness-behavior model.
Against: no attachment genetics.
The verdict is indirect.
CRP (rs1205, CRP).
For: C-reactive protein is the most practical inflammatory readout and tracks loneliness-related inflammation.
Against: CRP is a marker, not a behavioral gene, and the variant explains little behavioral variance.
The verdict is useful as a biomarker, weak as an attachment gene.
NFKB1 (rs28362491, NF-kB).
For: NF-kB is the master switch for the chronic social-stress inflammatory signal, the conserved transcriptional response to adversity.
Against: no direct attachment data.
The verdict is mechanistically central to the stress-inflammation link, empirically blank for attachment.
RELA (rs11820062, NF-kB complex).
For: RELA is a core NF-kB subunit, mechanistically inseparable from the inflammatory stress response.
Against: no attachment evidence.
The verdict is weak.
Methylation And One-Carbon Metabolism
This cluster does not encode bonding or threat at all, it encodes the machinery that writes and erases the epigenetic marks the rest of this map depends on, detailed in the methylation guide.
That makes it indirect but potentially upstream of everything.
MTHFR (rs1801133 C677T, rs1801131 A1298C, methylation capacity).
For: MTHFR sets the supply of methyl groups available to methylate genes like OXTR and NR3C1, so reduced MTHFR activity plausibly constrains the epigenetic regulation of attachment-relevant genes, covered in the MTHFR post.
Against: this is a general methylation-capacity argument with no study tying MTHFR genotype to attachment style, and MTHFR has been over-blamed for far more than it causes.
The verdict is upstream and biologically plausible, with no direct attachment evidence and a lot of hype to discount.
MTR (rs1805087, methylation cycle).
For: MTR regenerates methionine and so feeds the SAMe pool that all methylation draws on.
Against: no attachment link, purely upstream.
The verdict is a cofactor gene, not an attachment gene.
MTRR (rs1801394, B12 recycling).
For: MTRR keeps MTR running by recycling B12, sustaining methyl supply.
Against: no attachment data.
The verdict is upstream support only.
BHMT (rs3733890, methyl donor availability).
For: BHMT offers an alternative route to remethylate homocysteine and maintain methyl-donor availability.
Against: no attachment evidence.
The verdict is upstream support only.
CBS (rs234706, sulfur and methylation balance).
For: CBS pulls homocysteine into the transsulfuration pathway, shifting the balance between methylation and sulfur metabolism.
Against: no attachment link, and CBS is another gene heavily over-interpreted in consumer methylation circles.
The verdict is upstream and overhyped.
Sleep And Circadian Genes
This is the most emerging and most speculative cluster, included because circadian disruption degrades emotional regulation and relationship stability.
CLOCK (rs1801260, circadian rhythm).
For: the CLOCK 3111C variant has been tied to eveningness, mood instability, and sleep disruption, all of which erode the emotional regulation that relationships depend on.
Against: this is a mood-and-sleep correlate, with no attachment-style data and the usual candidate-SNP fragility.
The verdict is indirect and preliminary.
PER3 (VNTR, sleep regulation).
For: the PER3 VNTR shapes sleep timing and homeostatic sleep pressure, and poor sleep reliably worsens emotional reactivity.
Against: no attachment-specific evidence.
The verdict is indirect and preliminary.
The Overall Verdict

If you collapse the whole map, the genes sort into four honest tiers.
The strongest tier is epigenetic, not genetic, and it is small: OXTR methylation, NR3C1 methylation, and FKBP5 demethylation after early trauma each carry a real, mechanistically coherent, partly replicated link between early caregiving and adult relational biology. R R R
The second tier has a direct attachment or rejection finding but a fragile one: OPRM1 rejection sensitivity, AVPR1A pair bonding, COMT avoidant attachment, and DRD4 differential susceptibility each have a real published effect that is small, sex-dependent, or already dinged by a failed replication. R R R
The third tier is mechanistically convincing but measured on something other than attachment: FAAH and threat habituation, CD38 and oxytocin release, D2 and pair-bond reward, CRHR1 and trauma buffering, where the biology is strong and the attachment claim is inference. R R
The fourth and largest tier is pure inference, the GABA, glutamate, cholinergic, inflammation, sex-hormone, circadian, and most serotonin and norepinephrine entries, where the gene is plausibly in the circuit but has no human attachment data and inherits the candidate-gene replication problem wholesale. R R
The single most important conclusion is that attachment is not in your SNPs, it is in the methylation your early relationships wrote onto a handful of stress and bonding genes, and that mark is at least partly rewritable through the same relational and regulatory work that built it.
That is the optimistic reading of the whole field, and it is also the most evidence-based one.
Testing
You cannot test for an attachment style in a lab, and any company that claims to is selling a story.
What you can test is the underlying machinery, the methylation capacity, the stress-axis output, and the genotypes that bias the systems above.
Genetics
For the variants worth knowing, you can pull rs4680 (COMT), rs53576 (OXTR), rs1801133 and rs1801131 (MTHFR), rs6265 (BDNF), rs1360780 (FKBP5), and the rest from 23andMe or Ancestry raw data, which is the cheapest way to read your own genotypes.
I use the Methylation Genetics panel to read COMT, MTHFR, MTR, MTRR, and BHMT together, because the methylation cofactor genes determine whether any epigenetic mark can be written or maintained (Vibrant Wellness).
The honest caveat is that for most of the behavioral genes above, knowing your genotype changes very little, because the effects are small and environment-dependent.
Stress And Methylation Markers
Homocysteine is the practical readout of whether your methylation cycle has the methyl supply to maintain epigenetic marks, and elevation suggests the one-carbon machinery is constrained.
I use the Homocysteine with B12 and Folate panel to see methylation status and its cofactors in one draw (Quest Diagnostics).
The Cellular Zoomer covers organic acids and methylation markers, which shows how the catecholamine and methylation machinery is actually performing rather than just the genotype (Vibrant Wellness).
For the HPA-axis side, which is where the strongest attachment-epigenetics signal lives, a diurnal cortisol curve tells you whether the stress system that NR3C1 and FKBP5 regulate is actually dysregulated, and the Foundation Zoomer gives the surrounding metabolic, thyroid, and inflammatory context (Vibrant Wellness).
For interpreting any of this against your history and symptoms, this kind of bioindividual stress-system puzzle is exactly what a consult is built for.
How To Support The System
The first rule is the one this whole article has been building toward.
You cannot supplement your way to a secure attachment style, because attachment is a learned relational pattern and not a nutrient deficiency.
What you can do is support the physiology underneath it, the methylation machinery that maintains the epigenetic marks, the stress axis that attachment is fundamentally about, and the bonding chemistry that makes closeness register, and then do the relational and limbic work that actually reorganizes the pattern.
1. Maintain methylation capacity
The epigenetic marks on OXTR, NR3C1, and the rest are written and maintained with methyl groups, so cofactor status is upstream of the whole epigenetic layer.
- Methylfolate and methyl-B12: the central methyl-cycle inputs, dosed to your MTHFR status and started low if you are methylation-sensitive
- Magnesium glycinate: a cofactor across methylation and catecholamine clearance, and calming in its own right
- Pyridoxal-5-Phosphate (P5P): the active B6 needed across one-carbon and neurotransmitter metabolism
- Riboflavin: supports the MTHFR enzyme so methyl supply holds up
2. Regulate the stress axis
The strongest attachment-epigenetics signal is on the HPA axis, so calming a chronically activated stress system is the most leveraged physiological move, alongside the HPA-axis work any over-activated stress system needs.
- L-Theanine: raises alpha activity and takes the edge off sympathetic overarousal without sedation, covered in the theanine post
- Glycine: an inhibitory amino acid that is calming at night and feeds methylation buffering
- Magnesium L-threonate: crosses into the brain and supports NMDA modulation and stress recovery
- Taurine: membrane-stabilizing and dampens excitatory tone
3. Support bonding chemistry
These nudge the oxytocin signal that OXTR carries, and they are modest, preliminary, and no substitute for the bonding behavior that actually drives endogenous oxytocin.
- Vitamin D: vitamin D response elements sit on the oxytocin and oxytocin-receptor genes, so status may regulate oxytocin synthesis and response, detailed in the vitamin D cofactors post. R
- Magnesium: oxytocin receptors appear to need magnesium to function, increasing oxytocin's action at the receptor
- Vitamin C: a cofactor for the amidation enzyme that finishes oxytocin into its active form
4. Do the part supplements cannot
No capsule reorganizes an attachment strategy, and the interventions with the actual evidence base for changing attachment are relational and regulatory.
The list that does the real work is consistent, attuned relationships, fear-extinction and limbic work for the threat-driven patterns, sleep that stabilizes emotional regulation, and the slow repair of coregulation that was missing or broken early.
Lifestyle still matters as a floor, since stable sleep, sunlight, and the absence of chronic stressors keep the stress axis from overwriting any progress.
Mechanisms Of Action
Simple:
- Your early relationships left chemical tags on a few key stress and bonding genes, turning some of them up or down, and those tags, not your raw genes, are the closest thing to a biology of attachment.
- Most of the genes the internet links to attachment are only loosely connected, and the few that matter mostly work by shaping how strongly you feel threat and how rewarding closeness feels.
Advanced:
- Epigenetic programming of the stress axis early caregiving alters DNA methylation at the NR3C1 promoter and FKBP5 intronic regulatory regions, changing glucocorticoid-receptor expression and HPA-axis feedback sensitivity, which biases lifelong threat appraisal in close relationships. R R
- Oxytocin-receptor methylation as a bonding gain control OXTR promoter methylation sets receptor density, so higher methylation lowers oxytocin signaling and tracks attachment avoidance, while the rs53576 sequence variant explains little, making the epigenetic mark the operative variable. R R
- Oxytocin-dopamine coincidence detection pair-bond formation in the nucleus accumbens requires simultaneous oxytocin-receptor and dopamine D2-receptor activation, so the dopamine genes set how rewarding an oxytocin-tagged moment of closeness registers as. R
- Endocannabinoid tuning of threat habituation reduced FAAH activity raises anandamide, enhancing fronto-amygdala connectivity and fear extinction and blunting amygdala reactivity to threat, the felt-safety substrate that lets bonding cues land rather than trigger defense. R R
- Mu-opioid mediation of social pain the OPRM1 A118G variant modulates the social-pain response in dorsal anterior cingulate and insula, linking the opioid comfort system to rejection sensitivity, though the effect failed a large preregistered replication. R R
- Differential susceptibility as the unifying frame dopamine-related plasticity alleles like DRD4-7R and DAT1 increase environmental susceptibility rather than encoding a fixed deficit, so the same genotype produces worse or better attachment outcomes depending on caregiving quality. R R
More Research
Attachment genetics is best understood as a field that overpromised in its first decade and is still correcting, since the candidate-gene era produced famous findings like 5-HTTLPR and OPRM1 that did not survive large preregistered replication, which is the single most important context for reading any gene-and-attachment claim online. R R
Epigenetics is where the durable signal lives, because OXTR, NR3C1, and FKBP5 methylation give a mechanistic, partly replicated bridge from early caregiving to adult relational biology that no single SNP provides. R R
Differential susceptibility should replace the risk-gene framing entirely, since the genotypes that fare worst under harsh caregiving fare best under warm caregiving, which means the most useful question is not which alleles you carry but what environment they are being expressed in. R R
For biomarker and methylation testing I use the Methylation Genetics panel and the Cellular Zoomer together, since genotype plus functional methylation markers say more than either alone, and a diurnal cortisol curve adds the stress-axis output that the strongest attachment-epigenetics genes actually regulate.
Sex moderates a striking amount of this map, from the male-specific AVPR1A pair-bonding effect to the sex-specific CRHR1 buffering to the estrogen-by-oxytocin interactions, so any attachment-genetics interpretation that ignores sex is incomplete. R R
The most honest one-line summary is that there is no attachment gene, there is a small set of stress and bonding genes whose epigenetic settings your earliest relationships tuned, and the rest of the long list is mechanism in search of evidence.
Jacob Gordon
INHC, FMT-C
Board Certified Health Coach
I spent years battling unexplained chronic illness before discovering biohacking, epigenetics, and functional medicine. Now I share that research at MyBioHack to help others find their own answers.
Book a ConsultationRelated Protocols & Supplements
Deep-dive chapters and recommended supplements for this topic
Methylated B Complex
1 cap/day with food
SAMe
400mg on empty stomach
Resveratrol
250mg/day