Thymosin Peptides And Thymogens: The Thymus's Hormonal Language For Immune Regulation
By Jacob Gordon, INHC, FMT-C
The thymus spends the first two decades of your life building your T cell repertoire, then spends the rest slowly dismantling itself.
By age 40, roughly half of thymic tissue has been replaced by fat.
By age 70, functional thymic output has dropped to a fraction of its peak capacity.
What most people do not realize is that the thymus does not regulate immunity only by physically producing T cells.
It simultaneously secretes a family of hormone-like peptides that circulate in blood and act on immune cells throughout the body, functioning as a long-range endocrine system for immune calibration.
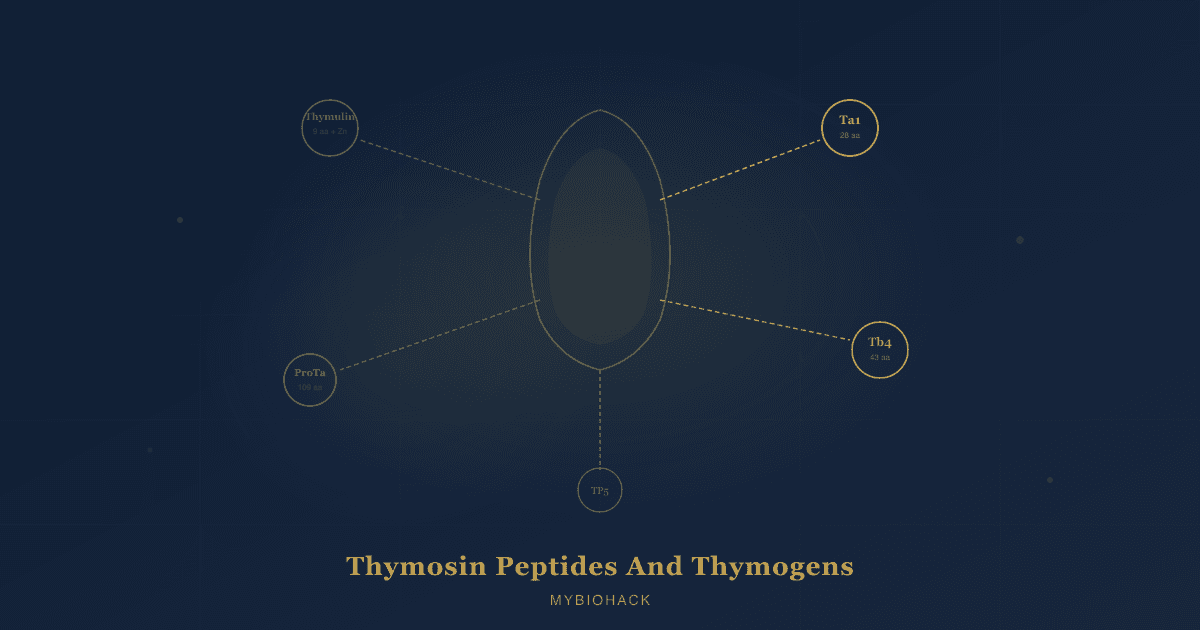
These peptides go by several names, including thymosin peptides, thymic hormones, and thymogens.
They are not a single molecule but a diverse family of structurally unrelated compounds with one common thread: they emerge from thymic tissue and exert measurable effects on immune cell differentiation, activation, and balance. R
The story starts with a crude extract in 1966 and has since unraveled into one of the most pharmacologically interesting immunology research programs of the past fifty years.
The Thymus As An Endocrine Organ
The thymus sits in the mediastinum behind the sternum and functions as the primary education center for T lymphocytes.
Bone marrow-derived T cell precursors migrate to the thymus, where they spend weeks undergoing positive selection (learning to recognize self-MHC) and negative selection (eliminating cells that would attack self-antigens). R
What is less widely appreciated is that the thymic epithelial cells (TECs) that orchestrate this education also secrete a collection of peptides into circulation.
These peptides reach peripheral lymphoid organs, circulating lymphocytes, and even the central nervous system, where they continue to regulate immune function far beyond the thymus itself. R
Thymic involution and the immunological consequences of losing this signaling:
Thymic involution begins around puberty and accelerates steadily with age.
As the gland atrophies, two things happen simultaneously: fewer naive T cells are exported to the periphery, and circulating levels of thymic peptide hormones decline. R
The immune system that results is one characterized by a contracted T cell repertoire, impaired response to novel antigens (including new pathogens and cancer neoantigens), a shift toward memory and effector cells rather than naive T cells, and chronic low-grade inflammation from dysregulated immune signaling. R
Zinc deficiency accelerates this process.
Thymulin, one of the primary thymic peptide hormones, requires zinc for biological activity.
In zinc-deficient animals and humans, thymulin activity drops and immune senescence accelerates in a pattern that partially mimics thymic atrophy. R
Zinc supplementation can restore thymulin activity in both young zinc-deficient and older zinc-adequate individuals whose zinc bioavailability has declined with age.
The thymus also maintains a bidirectional relationship with the hypothalamic-pituitary-adrenal (HPA) axis.
Corticosteroid levels (which rise with chronic stress and aging) promote thymic involution, while thymic peptides in turn influence neuroendocrine signaling.
Thymic hormone concentrations fluctuate with circadian rhythms in corticosteroid levels, and melatonin drives a nocturnal peak in thymosin alpha 1 and thymulin concentrations in both rats and humans. R
Quick Reference: Peptides At A Glance
The table below maps each thymic peptide to its primary roles, strongest clinical evidence, and the main reasons someone would or would not use it.
Detailed mechanisms follow in the individual sections.
| Peptide | Size | Primary Role | Strongest Evidence | Key Benefits | Downsides / Caveats |
|---|---|---|---|---|---|
| Thymosin Fraction 5 (TF5) | Mixed; >40 peptides | Broad T cell restoration; historical reference extract | Primary immunodeficiency; early cancer adjuvant trials | Broad immune restoration; foundational clinical rationale | Crude mixture; superseded by purified components; not reliably available |
| Thymosin Alpha 1 (Ta1 / Thymalfasin) | 28 aa | Innate and adaptive immune calibration via TLR3/4/9 | Chronic hepatitis B (approved 35+ countries); sepsis; HIV; vaccine adjuvant | Context-dependent immune balancing; activates where depressed, moderates where excessive; excellent safety record; Treg expansion; NK cell restoration | Subcutaneous injection only (no oral bioavailability); not FDA/EMA approved; COVID-19 mortality benefit inconclusive in meta-analysis; expensive via compounding |
| Prothymosin Alpha (ProTa) | 109 aa | Intracellular survival; extracellular TLR4-mediated innate activation | MS monocyte correction (in vitro); LAK/NK restoration in cancer patients | Broad immunostimulatory and adjuvant potential; antiviral; neuroprotective | Intracellular overexpression is a tumor proliferation marker; oncogenic risk of systemic use not resolved; no approved clinical formulation |
| Thymosin Beta 4 (Tb4) | 43 aa | G-actin sequestration; wound repair; angiogenesis; cardiac regeneration | Dry eye disease (RCT, Phase 2/3); wound healing (RCT); cardiac ischemia (Phase 2) | Potent wound closure and tissue regeneration; crosses blood-brain barrier; anti-fibrotic; anti-inflammatory; cardiac and neural repair | Dual role in cancer: same angiogenic/migratory properties that heal tissue can promote tumor invasion; no oral bioavailability; not approved (compassionate/investigational only); short plasma half-life |
| Thymosin Beta 10 (Tb10) | 43 aa | G-actin sequestration; muscle regeneration | Mostly preclinical; skeletal muscle repair models | Similar repair biology to Tb4; early chemotaxis data | Overexpressed in several cancers (thyroid, ovarian, pancreatic); primarily a research compound; no clinical development program |
| Thymosin Beta 15 (Tb15) | 43 aa | G-actin sequestration; cell motility | Prostate cancer biomarker research | No therapeutic benefit established | Primarily associated with tumor invasion and poor prognosis in prostate cancer; no therapeutic use |
| Thymulin | 9 aa (nonapeptide) | T cell precursor maturation; requires zinc for activity | Zinc-dependent thymic output; animal models of infection and aging | Direct thymic hormone; activity restorable with zinc supplementation; anti-inflammatory properties; analgesic in CNS models | Levels near undetectable by age 60+; zinc deficiency abrogates activity even when peptide levels are normal; no approved clinical formulation; distinguishing peptide vs zinc deficiency requires specialized testing |
| Thymopoietin | 49 aa | T cell precursor differentiation from bone marrow | Animal models of infection and aging; antigen presentation via MHC II | Drives thymic seeding; modulates T cell cytokine profiles | High levels associated with worse cancer outcomes (gastric, pancreatic); tumor-promoting properties limit therapeutic use; not in clinical development |
| Thymopentin (TP5) | 5 aa (pentapeptide) | T cell maturation; HLA-DR-mediated APC signaling | Approved in China (immunodeficiencies, RA, AIDS, hepatitis B); recent tumor immunology data (T cell exhaustion reversal) | Active site of thymopoietin; simpler structure; approved clinical experience; recent thymic rejuvenation and T cell exhaustion data promising | Plasma half-life <30 seconds; no oral bioavailability; injection required; sustained-release formulations still investigational; not FDA/EMA approved |
| Thymic Humoral Factor (THF) | Variable | Hematopoietic progenitor stimulation; antiviral immune response | Antiviral cytokine induction in chronic HBV (in vitro); bone marrow stimulation | Unique hematopoietic progenitor effects | Not in clinical development; primarily historical and mechanistic interest; limited modern research |
Reading the table:
If immune restoration in infection, post-chemotherapy, or aging is the goal, Ta1 is the starting point.
It has the deepest clinical evidence base, the best safety record, and the most validated mechanism.
If tissue repair, wound healing, corneal injury, or cardiac regeneration is the goal, Tb4 is the leading candidate, with the caveat that active malignancy requires careful consideration of its angiogenic properties.
If thymic hormone replacement per se is the conceptual goal (especially in aging), thymulin points toward zinc optimization as the most accessible intervention before any peptide supplementation.
Many people presenting with apparent thymulin deficiency have zinc deficiency as the primary driver.
Tb10, Tb15, thymopoietin, and THF are primarily research compounds and are not therapeutic candidates for the clinical contexts that bring most readers to this page.
Thymopoietin's tumor-promoting associations specifically argue against unsupervised supplementation.
Thymosin Fraction 5: Where It All Started
What it is: A partially purified, heat-stable preparation of thymic peptides from bovine thymus tissue
In 1966, Abraham White and Allan Goldstein at the Albert Einstein College of Medicine isolated a preparation from calf thymus that could restore immune function in thymectomized mice.
They named it thymosin, and after further purification developed a standardized extract they called thymosin fraction 5 (TF5). R
TF5 is not a single molecule.
It is a complex mixture of at least 10 major polypeptides and more than 30 additional minor peptides, with molecular weights mostly below 15,000 Da.
These include polypeptide beta-1, thymosin alpha 1, prothymosin alpha, parathymosin, and multiple beta-thymosins, all acting individually, sequentially, or in concert. R
What TF5 does:
TF5 stimulates T cell differentiation and maturation, enhances immune function in immunocompromised hosts, induces cytotoxic T cell development in mixed lymphocyte tumor responses, and demonstrates anti-tumor potential in experimental models through T cell-mediated mechanisms. R
In early clinical trials with primary immunodeficiency diseases, cancer, and autoimmune conditions, TF5 demonstrated immunorestorative properties that supported the concept of thymic endocrine function and motivated the effort to isolate and characterize individual active peptides.
The isolation cascade:
TF5's promising effects in the 1970s drove researchers to fractionate it further.
This led to the isolation of thymosin alpha 1 in 1977, which proved to be 10 to 1000 times more active than TF5 in standardized bioassays. R
The subsequent decades produced characterization of thymosin beta 4, prothymosin alpha, and other family members, each with distinct mechanisms.
Current status:
TF5 itself is rarely used clinically today, having been largely superseded by its purified components, particularly thymosin alpha 1.
Its legacy is foundational: it established the concept of thymic endocrine function and provided the biological rationale for all subsequent thymosin-based therapeutic development.
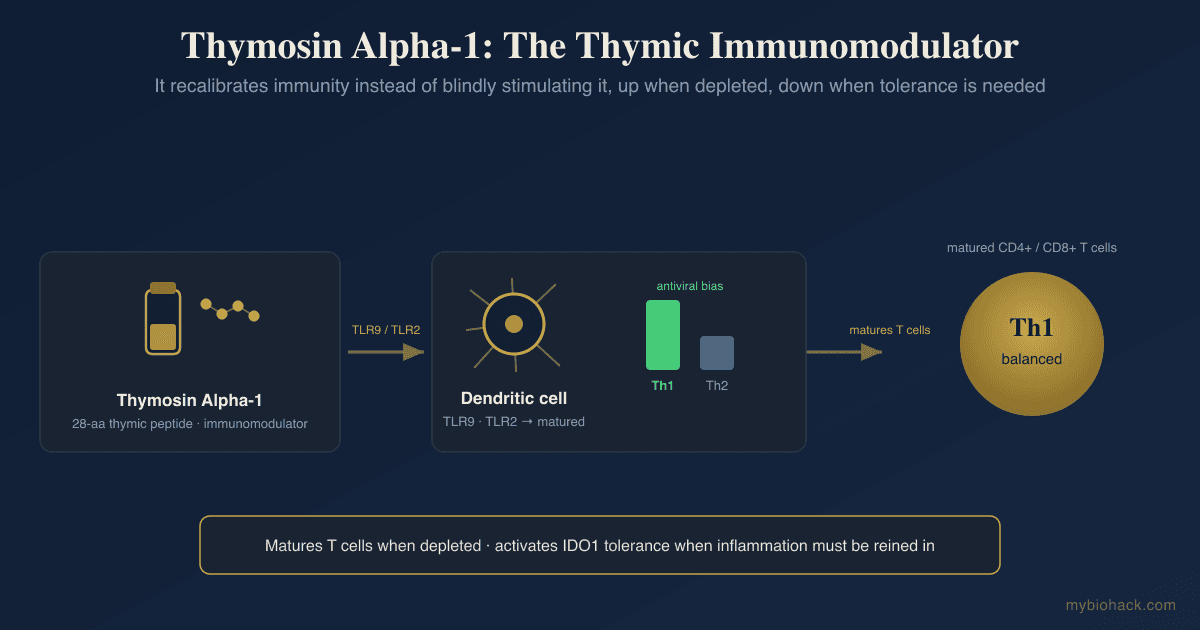
Thymosin Alpha 1 (Ta1)
What it is: 28-amino acid peptide; N-terminal cleavage product of prothymosin alpha; the most clinically advanced thymic peptide
Sequence: Ac-Ser-Asp-Ala-Ala-Val-Asp-Thr-Ser-Ser-Glu-Ile-Thr-Thr-Lys-Asp-Leu-Lys-Glu-Lys-Lys-Glu-Val-Val-Glu-Glu-Ala-Glu-Asn-OH
Molecular weight: Approximately 3,108 Da
Synthetic analog (approved drug): Thymalfasin (Zadaxin)
Thymosin alpha 1 (Ta1) is produced by asparagine endopeptidase (legumain) cleavage of prothymosin alpha at the N-terminal end. R
It circulates at detectable levels in healthy individuals, with the highest concentrations in lymphoid tissues.
Levels decline with age and are altered in immunodeficiency states.
Females have lower serum Ta1 than males in healthy populations, a difference that may contribute to sex-based differences in immune function and autoimmune disease susceptibility. R
Mechanism Of Action
Context-dependent immunomodulation:
Ta1 does not simply activate or suppress immunity.
Its effects are context-dependent, meaning it restores immune balance rather than pushing uniformly in one direction.
In immunodeficient hosts, Ta1 is immune-activating; in states of excessive immune activation, it can be immune-moderating. R
TLR-mediated innate immune priming:
Ta1 acts through toll-like receptors (TLRs) on both myeloid and plasmacytoid dendritic cells (pDCs).
It binds TLR3, TLR4, and TLR9, activating downstream IRF3 and NF-κB signaling pathways to initiate cytokine production.
TLR2 and TLR7 also contribute to Ta1 signaling in some contexts. R
This TLR-mediated innate priming is why Ta1 can function as a vaccine adjuvant and why its immune-activating effects begin with the innate system before propagating to adaptive T and B cell responses.
Dendritic cell maturation:
Ta1 matures dendritic cells (DCs), enhancing their antigen-presenting capacity.
In myeloid DCs, Ta1 drives maturation and Th1 polarization.
In pDCs, Ta1 promotes IFN-α production critical for antiviral responses. R
T cell effects:
Ta1 enhances the expression of IL-2 receptors on T cells and promotes IL-2 production and internalization, supporting T cell proliferation and differentiation.
It enhances CD4+ helper T cell function and has a documented Th1-polarizing effect (promoting IFN-γ, IL-2, IL-12, IL-18), which improves antiviral and antifungal responses. R
It also expands regulatory T cell (Treg) populations, which is the likely mechanism behind its immune-moderating effect in conditions of autoimmune or inflammatory excess. R
IDO-dependent immune modulation:
Ta1 can attenuate the immunogenic activity of myeloid DCs through an indoleamine 2,3-dioxygenase (IDO)-dependent pathway.
IDO converts tryptophan to kynurenine, shifting the local immune environment toward tolerance.
This dual capacity (activating innate and adaptive immunity while simultaneously engaging tolerogenic pathways via IDO) qualifies Ta1 as a genuinely pleiotropic immunomodulatory peptide capable of fine-tuning immune quality rather than merely amplifying immune magnitude. R
NK cell activity:
Ta1 restores depressed natural killer (NK) cell and lymphokine-activated killer (LAK) cell activity in immunosuppressed patients.
It also partially reverses steroid-induced apoptosis of thymocytes, relevant in patients receiving glucocorticoid therapy. R
Clinical Evidence
Chronic hepatitis B:
This is the strongest evidence base for Ta1.
Thymalfasin is approved for hepatitis B treatment in more than 35 countries (though not the US or EU).
In randomized controlled trials of chronic hepatitis B, Ta1 combined with interferon significantly improved rates of HBeAg seroconversion and HBV DNA suppression compared to interferon alone. R
The mechanism involves both direct antiviral immune priming via TLR9 and restoration of the exhausted T cell and NK cell responses characteristic of chronic HBV infection.
Chronic hepatitis C:
Ta1 has been studied as an add-on to standard-of-care interferon plus ribavirin therapy in hepatitis C, with some evidence of improved response rates particularly in non-responders and in patients coinfected with HIV.
HIV:
Ta1 increases CD4+ T cell counts, promotes IL-2 receptor expression, reduces viral load in some studies, and has been shown to increase T cell receptor excision circles (TRECs), a marker of thymic output.
The safety and tolerability profile is well established in HIV patients, and high-dose prolonged use appears more effective than short courses. R
Sepsis:
Several randomized controlled trials in severe sepsis have evaluated Ta1, motivated by its ability to restore immune function in profoundly immunocompromised septic patients who exhibit T cell exhaustion, lymphopenia, and DC dysfunction.
Meta-analyses suggest Ta1 reduces 28-day mortality in sepsis, though individual trial results have been mixed. R
COVID-19:
Ta1 received significant attention during COVID-19 because the immunological profile of severe COVID-19 (lymphopenia, T cell exhaustion, NK cell dysfunction, dysregulated innate-to-adaptive transition) closely matches the conditions where Ta1 has demonstrated benefit.
Early observational and small interventional studies suggested reduced mortality in severe disease.
A subsequent meta-analysis found no significant mortality benefit, though methodological heterogeneity (variable dosing, timing, patient selection) limits interpretation. R
Cancer:
Ta1 has been studied as an adjunct to standard cancer therapy, with the strongest evidence in hepatocellular carcinoma, non-small cell lung cancer, and melanoma.
It improves immune cell counts and function in chemotherapy- and radiotherapy-induced immunosuppression.
More recently, retrospective data from non-small cell lung cancer patients undergoing chemoradiotherapy plus immune checkpoint inhibitors showed that those who also received Ta1 had better outcomes, with suggestions of reduced rates of pneumonitis and improved immune recovery from lymphopenia. R
Vaccine adjuvant:
Ta1 is among the most studied thymic peptide vaccine adjuvants.
It enhances antibody titers and cellular immune responses to influenza, hepatitis B, and other vaccines, particularly in elderly or immunocompromised individuals with blunted baseline responses.
The TLR3/4/9 agonism mechanism explains why Ta1 improves innate immune priming for adaptive vaccine response.
Autoimmune disease:
Ta1 serum levels are lower in patients with rheumatoid arthritis, psoriatic arthritis, and SLE compared to healthy controls. R
The Treg-expanding and IDO-mediated tolerogenic properties of Ta1 make it theoretically useful in autoimmunity, though clinical evidence is limited relative to the infectious disease and oncology data.
Antifungal infections:
Ta1 enhances antifungal defenses by priming dendritic cells and expanding Th1 and Treg populations, generating IFN-γ-driven macrophage activation and cytotoxic T cell responses against fungal pathogens.
This has been demonstrated in invasive aspergillosis and Candida infections in immunocompromised hosts. R
Administration And Safety
Ta1 is administered subcutaneously or intramuscularly.
Standard dosing in hepatitis B studies uses 1.6 mg twice weekly for 6 to 12 months.
There is no established oral bioavailability; oral administration is not effective.
The safety profile across decades of clinical use is excellent: no serious adverse effects have been reported in any major trial.
Minor injection site reactions occur rarely. R
Prothymosin Alpha (ProTa)
What it is: 109 to 112 amino acid polypeptide; intracellular precursor to thymosin alpha 1
Prothymosin alpha (ProTa) is the intact molecular precursor from which thymosin alpha 1 is cleaved.
It is a highly acidic, intrinsically disordered protein (no stable tertiary structure under physiological conditions) that is ubiquitously expressed in virtually all mammalian tissues, not just the thymus. R
Dual role by localization:
ProTa has a distinct dual biological identity depending on its cellular location.
Intracellularly, ProTa functions as a survival and proliferation mediator.
It interacts with chromatin remodeling complexes, promotes cell cycle progression, and protects cells from apoptosis.
High intracellular ProTa expression is therefore associated with proliferating cells including cancer cells, and ProTa overexpression has been identified as a potential tumor biomarker. R
Extracellularly, ProTa functions as an alarmin (damage-associated molecular pattern) and biological response modifier.
When released from necrotic or stressed cells, extracellular ProTa signals through TLR4 on dendritic cells and macrophages, triggering innate immune activation.
It enhances NK cell cytolytic activity, restores depressed allogeneic cell-mediated cytotoxicity in cancer patients, and promotes LAK cell induction. R
The C-terminal decapeptide ProTa(100-109):
The primary immunoactive region of ProTa is the C-terminal decapeptide ProTa(100-109), which retains much of the full-length molecule's adjuvant and immunostimulatory activity while being far smaller.
ProTa(100-109) and the full-length molecule both signal through TLR4 to activate dendritic cells and has been proposed as a candidate cancer immunotherapy adjuvant. R
Multiple sclerosis:
Purified ProTa has been shown to regulate the defective monocyte phenotype observed in multiple sclerosis (MS), improving antigen presentation and T cell activation.
ProTa restores abnormal monocyte-T cell signaling in MS patients in vitro, suggesting therapeutic potential for autoimmune neurological disease. R
Cancer diagnostic marker:
Because ProTa is highly expressed in proliferating cells, elevated ProTa levels in tumor tissue or serum may serve as a prognostic marker in certain cancers.
High ProTa expression has been observed in hepatocellular carcinoma, colorectal cancer, and renal cell carcinoma. R
Whether ProTa levels will translate to a validated clinical biomarker remains an active area.
Thymosin Beta 4 (Tb4)
What it is: 43-amino acid acidic peptide (pI 4.6); most abundant intracellular actin-sequestering protein in most mammalian cells
Molecular weight: Approximately 4,963 Da
Thymosin beta 4 (Tb4) was isolated from thymosin fraction 5 but has since been found to be the most abundant and ubiquitously distributed of all thymosin peptides.
It is present in essentially all nucleated mammalian cells at high concentrations, with particularly high levels in blood platelets, neutrophils, macrophages, and other lymphoid tissues. R
Despite its original isolation from thymus, Tb4 is not thymus-specific and is better understood as a general tissue repair and cytoskeletal regulation peptide that happens to also modulate immune function.
Mechanism Of Action
G-actin sequestration and cytoskeletal dynamics:
The primary intracellular function of Tb4 is to bind and sequester G-actin (globular, monomeric actin), keeping it in an unpolymerized state available for rapid actin filament (F-actin) nucleation when needed.
This regulates actin cytoskeletal dynamics throughout the cell, controlling cell motility, shape changes, endocytosis, and organogenesis. R
By controlling the available pool of G-actin, Tb4 directly influences virtually every cellular process that requires cytoskeletal rearrangement, including cell migration, division, differentiation, and wound closure.
Extracellular functions:
After injury, Tb4 is actively released by platelets, macrophages, and other cell types to function as an extracellular paracrine and autocrine signal. R
Once outside the cell, Tb4 promotes:
- Keratinocyte and fibroblast migration and wound closure
- Angiogenesis via VEGF, angiopoietin 2, and Notch-VEGF signaling
- Stem and progenitor cell mobilization, migration, and differentiation
- Reduction of myofibroblast differentiation (reducing scar formation and fibrosis)
- Anti-apoptotic signaling via ILK (integrin-linked kinase) pathway activation
Cardiac and vascular regeneration:
Tb4's cardiovascular biology is one of its most studied dimensions.
Tb4 is expressed in the developing heart and plays essential roles in all key stages of cardiac vessel development: vasculogenesis, angiogenesis, and arteriogenesis. R
After myocardial infarction, Tb4 promotes cardiomyocyte survival in hypoxia, stimulates neoangiogenesis in the infarcted territory, and activates resident epicardial progenitor cells to regenerate cardiac tissue.
In combination with cardiac transcription factors (Gata4, Mef2c, Tbx5), Tb4 synergistically enhances cardiac fibroblast reprogramming to cardiomyocyte-like cells, a potential regenerative strategy. R
Anti-inflammatory effects:
Tb4 reduces the production of inflammatory cytokines and chemokines in multiple injury models.
It inhibits activated hepatic stellate cell proliferation and attenuates liver fibrosis via Notch signaling inhibition. R
In ulcerative colitis models, Tb4 reduces intestinal inflammation and promotes epithelial repair.
In the brain, Tb4 can cross the blood-brain barrier and acts on multiple neural cell types: it provides neuroprotection, immunosuppression of microglial hyperactivation, and neurorestoration including remyelination, synaptogenesis, and axon growth. R
Muscle regeneration:
After skeletal muscle injury, Tb4 mRNA is upregulated in both regenerating muscle fibers and infiltrating inflammatory hematopoietic cells.
Tb4 and its sulphoxidized form (Tb4 sulfoxide) accelerate wound closure and function as chemoattractants for satellite cell-derived myoblasts, facilitating muscle repair. R
Clinical Evidence
Wound healing:
Tb4 was studied in randomized controlled trials for chronic pressure ulcers and stasis ulcers (as topical RGN-352 or systemic TB4).
In one early trial, topical Tb4 significantly accelerated wound closure and reepithelialization compared to placebo.
In full-thickness wound models, Tb4 increased collagen deposition and angiogenesis by 42 to 61% within the first week. R
Dry eye disease (xerophthalmia):
Tb4-containing eye drops (RGN-259, developed by RegeneRx) have been studied in randomized phase 2 and phase 3 trials for moderate-to-severe dry eye disease.
The compound promotes corneal epithelial repair and reduces ocular surface inflammation.
Phase 2 results were positive; larger phase 3 development is ongoing. R
Ischemic heart disease:
Tb4 has been in development as an intravenous treatment for acute myocardial infarction (PREMISE trial by RegeneRx), based on its cardiac survival, angiogenesis, and progenitor cell activation properties.
Early phase results in acute MI demonstrated safety and signals of cardiac preservation.
Multiple sclerosis and neurological injury:
Tb4's ability to cross the blood-brain barrier and promote remyelination, oligodendrocyte survival, and neurovascular remodeling makes it a candidate for MS treatment.
Preclinical data in EAE (experimental autoimmune encephalomyelitis) and stroke models are favorable.
Clinical translation in MS is in early stages. R
Fibrosis:
Tb4 has demonstrated anti-fibrotic effects across pulmonary, hepatic, renal, and cardiac fibrosis models by reducing myofibroblast activation and TGF-β-driven extracellular matrix accumulation.
Administration:
Tb4 can be administered systemically (intravenous, subcutaneous), topically (eye drops, wound gels), or via local injection.
It crosses the blood-brain barrier.
Half-life after systemic administration is short (minutes), requiring either repeated dosing or sustained-release formulations.
Safety data across trials is consistently favorable.
The cancer ambiguity:
Tb4 presents a complex picture in oncology.
Its promotion of cell migration, angiogenesis, and survival pathways that are beneficial in tissue repair are also characteristics that can facilitate tumor progression and metastasis when hijacked by cancer cells.
Elevated Tb4 is associated with invasion and poor prognosis in some cancers.
In other settings, Tb4 has shown tumor-suppressive effects.
This duality is context-dependent and tumor-type-specific and should be taken into account in oncology contexts. R
Thymosin Beta 10 And Beta 15
Thymosin beta 10 (Tb10):
Tb10 is structurally related to Tb4 and shares its G-actin-sequestering function.
It is upregulated during early skeletal muscle regeneration (alongside Tb4) and promotes myoblast chemotaxis in a manner similar to Tb4 sulfoxide. R
Tb10 is also overexpressed in several cancers (including thyroid cancer, ovarian cancer, and pancreatic cancer) and has been proposed as a tumor progression marker.
Its role in tumorigenesis via actin cytoskeletal regulation and cell motility promotion parallels the Tb4 cancer ambiguity. R
Notably, despite being widely overexpressed in tumor tissue, Tb10 (like Tb4) has demonstrated suppressive tumor effects in some disease contexts, underscoring that the beta-thymosin family's role in cancer biology is context- and tumor-type-dependent.
Thymosin beta 15 (Tb15):
Tb15 is the least studied of the beta-thymosin family in terms of therapeutic development.
It is overexpressed in aggressive prostate cancer and has been studied as a potential biomarker for disease progression and metastatic potential.
Tb15 promotes cell motility via actin cytoskeletal regulation similarly to Tb4, but unlike Tb4, it has not demonstrated clear tissue repair or regenerative benefits in the same way.
The available evidence associates Tb15 most strongly with oncogenic processes rather than therapeutic applications. R
Thymulin: The Zinc-Dependent Thymic Hormone
What it is: Nonapeptide (9 amino acids); requires zinc ion for biological activity; produced exclusively by thymic epithelial cells
Also known as: Facteur thymique serique (FTS), serum thymic factor (STF)
Thymulin (TH) was first isolated by the team of Professor J.F. Bach in 1975 from pig serum, where it circulates at low but measurable concentrations as a true endocrine hormone.
It is the only confirmed thymus-specific peptide hormone in the sense that thymic epithelial cells appear to be its exclusive source. R
The Zinc Requirement
Thymulin is structurally unique among thymic peptides because it is a metallopeptide.
Biological activity depends absolutely on zinc ion conjugation.
Zinc-free thymulin (apoThymulin) is biologically inactive; only zinc-bound thymulin (ZnThymulin) activates T cell-related pathways. R
This creates a direct mechanistic link between dietary zinc status and thymic immune output.
In zinc-deficient animals, thymulin activity is suppressed even when thymulin protein levels are normal.
Zinc supplementation restores thymulin activity, explaining part of the well-documented immune-restorative effect of zinc supplementation in zinc-deficient and elderly individuals.
The clinical implication: measuring total thymulin versus biologically active thymulin (requiring zinc bioavailability measurement) can reveal whether apparent thymulin deficiency is truly peptide deficiency or zinc deficiency masquerading as thymulin insufficiency.
Blood Levels And Age-Related Decline
Thymulin concentrations in blood peak in the early postnatal period (approximately 2 pg/mL in umbilical vessels) and decline progressively with age.
By the sixth decade, biologically active thymulin in circulation is near or below detectable limits in most individuals, correlating with the age-related decline in cell-mediated immunity. R
Melatonin increases nighttime thymulin secretion, and thymulin and thymosin alpha 1 follow parallel nocturnal peaks driven by melatonin rhythms. R
Disruption of circadian rhythms, including shift work and artificial light exposure at night, likely suppresses this thymulin surge.
Mechanism And Immunological Effects
Thymulin acts on T cell precursors within and immediately outside the thymus, promoting acquisition of T cell surface markers and functional maturation.
In thymectomized animals, thymulin partially restores T cell function, the original defining property of a thymic hormone. R
Thymulin modulates cytokine production in a context-dependent way:
- In healthy donors, thymulin increases IFN-γ, IL-2, and IL-1α production by mitogen-stimulated cells.
- In SLE patients, thymulin inhibits these same cytokines (IFN-γ, IL-2, IL-1α, and also IL-6 and TNF-α), demonstrating its immunonormalizing rather than uniformly activating role. R
Thymulin and thymopentin together restore antibody avidity in aged or thymectomized animals, enhance antibody production in aging mice, and support delayed-type hypersensitivity responses.
Anti-inflammatory and analgesic properties:
Beyond immune modulation, thymulin demonstrates anti-inflammatory and analgesic properties in the central nervous system.
These nociceptive effects have been documented in animal models but have not entered clinical trials. R
Cancer:
Limited data suggests thymulin promotes anti-tumor effects, consistent with its role in maintaining cell-mediated immunity.
It has not been developed as a clinical cancer therapeutic to the extent that Ta1 has.
Thymopoietin And Thymopentin (TP5)
Thymopoietin
What it is: 49-amino acid peptide (5.5 kDa); three isoforms: thymopoietin alpha (75 kDa), thymopoietin beta (51 kDa, also known as laminin-associated polypeptide 2), thymopoietin gamma; all secreted by thymic epithelial cells
Thymopoietin was originally identified as the factor responsible for inducing T cell differentiation from bone marrow precursors.
Its three isoforms share the same 49-amino acid region but differ in N-terminal extensions. R
Thymopoietin's immunoregulatory actions on peripheral T cells are mediated via intracellular cyclic GMP (cGMP) elevations (in contrast to the cyclic AMP elevations induced in precursor T cells that drive T cell differentiation). R
This distinguishes the mechanism of thymopoietin on mature versus immature T cells.
Thymopoietin directly binds to antigen-presenting cells at MHC class II molecules, which may mediate part of its T cell modulatory effect. R
The cancer picture for thymopoietin is problematic: high thymopoietin levels have been associated with significantly poorer overall survival in gastric cancer, and reducing thymopoietin levels in pancreatic cancer inhibited cell proliferation, migration, and invasion.
The isoform lamin-associated polypeptide 2 (thymopoietin beta) is a nuclear structural protein expressed in virtually all proliferating cells.
These observations suggest thymopoietin has tumor-promoting properties that likely distinguish it from the other thymic peptides in oncology contexts. R
Thymopentin (TP5)
What it is: Synthetic pentapeptide corresponding to amino acids 32 to 36 of thymopoietin; sequence: Arg-Lys-Asp-Val-Tyr
Molecular weight: 679.77 Da
Trade name/approval: Approved as an immunomodulator in China; previously studied in the US and Europe
Thymopentin was identified as the active site of thymopoietin after it was demonstrated that residues 32 to 36 (RKDVY) reproduce all the biological activity of the parent molecule. R
It is substantially simpler to synthesize than the full 49-amino acid thymopoietin, making it more practical as a therapeutic.
Mechanism:
TP5 promotes differentiation of thymocytes and modulates function of mature T cells.
It binds to HLA-DR molecules on antigen-presenting cells (APCs), suggesting that its T cell modulatory effect is mediated through antigen presentation rather than direct T cell receptor engagement. R
In peripheral T cells, TP5 modulates immune responses via cGMP (as does full-length thymopoietin), increasing IL-2 and IFN-γ but reducing IL-4, shifting toward Th1 and away from Th2 immune profiles.
Bone marrow T lymphocyte precursors treated with TP5 show increased thymus colonization, suggesting TP5 may enhance thymic seeding and T cell production in addition to its peripheral effects. R
Recent tumor immunology data:
A 2025 study demonstrated that TP5 potently suppresses tumor growth across multiple murine cancer models through strictly T cell-dependent mechanisms.
TP5 promoted thymic rejuvenation under immunocompromised conditions, restoring thymus-tumor immunological balance and revitalizing peripheral T cell immunity.
It functionally reprogrammed T cell states, preserving effector function while ameliorating exhaustion. R
In T cell adoptive therapy models, TP5-expanded T cells showed enhanced proliferation and effector function after transfer.
The half-life problem:
TP5's primary clinical limitation is an extremely short plasma half-life, measured at less than 30 seconds after intravenous injection.
It is extensively metabolized in the gastrointestinal tract, making oral bioavailability negligible.
Standard clinical use requires subcutaneous or intramuscular injection, and the short half-life has driven formulation research into sustained-release systems.
A phospholipid-based phase separation gel (PPSG) formulation has demonstrated that a single injection of TP5-PPSG provides an immunoregulatory effect equivalent to 14 consecutive daily TP5 injections in immunosuppressed rats. R
Clinical applications:
TP5 has been used clinically for chronic lymphocytic leukemia, rheumatoid arthritis, AIDS, cancer immunodeficiency, and chronic hepatitis B.
In animal models, TP5 significantly increases resistance to Leishmania major infection in aged mice, consistent with enhanced innate and adaptive immune function.
In severe burns and bacterial infection models, TP5 administration increased survival rates from 40 to 80%. R
Thymic Humoral Factor (THF)
What it is: A family of peptides isolated from thymic tissue and serum; less structurally defined than the other thymic hormones
Thymic humoral factor (THF) refers to a group of thymus-derived peptides that were isolated and characterized starting in the 1970s, primarily through the work of Nathan Trainin's laboratory.
THF-gamma 2 (THFγ2) is the best characterized, a short peptide with documented effects on hematopoietic progenitor cell proliferation and viral infection responses.
Peripheral blood mononuclear cells from patients with chronic hepatitis B infection produce increased IL-2 and TNF-α in response to THFγ2, suggesting an antiviral immunostimulatory role.
THFγ2 has also increased the number of myeloid colonies from bone marrow cells, and human erythroid and myeloid hematopoietic progenitor cells are stimulated to grow by THFγ2. R
THF preparations were at one point studied in clinical contexts (including as TFX in Poland), with immunomodulatory effects demonstrated in animal models, but THF preparations never advanced to major clinical trials and are now primarily of historical and mechanistic research interest. R
Clinical Applications By Condition
Immunodeficiency And Thymic Failure
Primary immunodeficiencies:
Ta1 and TF5 have the strongest track record in restoring immune function in primary T cell deficiencies.
Thymic peptides partially compensate for absent or hypofunctional thymic tissue by supplying the hormonal signals that normally drive peripheral T cell differentiation and activation.
Post-chemotherapy and radiation immune suppression:
Ta1 is among the best-supported approaches for accelerating immune recovery after chemotherapy-induced lymphopenia.
Its TLR-mediated innate priming and T cell receptor upregulation restore immune competence more quickly than recovery alone.
Aging-associated immunosenescence:
The decline in thymulin, Ta1, and other thymic peptides with age mirrors the decline in immune competence.
Zinc supplementation to restore thymulin bioactivity, combined with Ta1 supplementation, addresses the most tractable hormonal components of immunosenescence.
Both have safety profiles that support use in older adults.
Viral Infections
The mechanism is consistent across viruses: thymic peptides (particularly Ta1 via TLR3/4/9) restore the innate immune priming of dendritic cells, normalize T cell function, and enhance NK cell activity that is suppressed during chronic viral infection.
Hepatitis B: Ta1 is approved in multiple countries; the most robust clinical evidence.
Hepatitis C: Ta1 as adjuvant to interferon-based regimens.
HIV: Ta1 increases CD4+ counts, thymic output (TRECs), and reduces viral load in some studies.
COVID-19 and post-viral syndromes: Evidence base developing; strongest signals in severe disease with lymphopenia.
Wound Healing And Tissue Repair
Tb4 is the dominant peptide for this application.
Its actin-sequestration mechanism drives keratinocyte migration, fibroblast activation, angiogenesis, and the full wound repair cascade.
Corneal injury (dry eye disease trials) and chronic wound healing are the most advanced clinical development areas.
Cardiac regeneration after myocardial infarction is the highest-stakes potential application.
Cancer Immunotherapy Adjuvant
Ta1 has the strongest evidence as a cancer immunotherapy adjuvant, primarily by restoring immune competence in the immunosuppressed cancer patient.
TP5 is showing early promise in tumor immunology through thymic rejuvenation and T cell exhaustion reversal.
Tb4's role in cancer is complicated by its dual tissue-repair/tumor-progression biology and should be approached with caution in active malignancy without better understanding of tumor-specific context.
Autoimmune Disease
Ta1's IDO-mediated tolerogenic pathway and Treg expansion capacity make it theoretically useful in autoimmunity.
Its demonstrated serum deficit in rheumatoid arthritis, psoriatic arthritis, and SLE patients supports a mechanistic case.
TP5 and thymulin modulate cytokines in SLE-specific ways (suppressing rather than activating in the SLE context).
None of these peptides have robust large-scale RCT data in autoimmune disease comparable to the viral infection data.
Testing And Optimization
Thymic peptide blood tests:
No routinely available clinical lab panels directly measure thymosin alpha 1, thymulin, or Tb4.
Thymulin can be measured in research settings as total vs. active (zinc-bound) thymulin using bioassays; this distinction matters clinically because zinc deficiency suppresses activity without reducing peptide levels.
ProTa levels in tissue are studied as a cancer biomarker but are not part of routine panels.
Functional markers relevant to thymic output:
T-cell receptor excision circles (TRECs) in peripheral blood are the most direct clinical measure of thymic output, reflecting recently exported naive T cells.
Low TRECs indicate reduced thymic output and are measurable in specialized immunology labs.
CD4/CD8 ratio and naive versus memory T cell proportions on lymphocyte subset panels reflect the cumulative consequence of thymic function over time.
Zinc:
Given thymulin's zinc requirement, zinc status is functionally a proxy for part of thymulin activity.
Serum zinc, red blood cell zinc, or urinary zinc can be assessed.
Zinc is included in many micronutrient panels.
Immune panels that inform thymic peptide status:
A comprehensive lymphocyte subset panel measuring naive T cells (CD45RA+CCR7+ within CD4+ and CD8+ compartments), recent thymic emigrants (CD31+ naive T cells), Tregs (CD4+CD25+FoxP3+), and NK cell numbers and activity provides the best functional picture of what thymic peptide activity is actually delivering to peripheral immunity.
The Cellular Zoomer panel is the most accessible starting point for understanding lymphocyte compartment health.
Practical optimization stack:
Zinc supplementation (15 to 30 mg elemental zinc) is the most accessible intervention that directly supports thymulin bioactivity.
Ta1 (thymalfasin) is available by injection through compounding pharmacies in the US for off-label use.
Standard dose is 1.6 mg subcutaneously twice weekly to daily depending on clinical context.
Tb4 eye drops (RGN-259) are in late clinical trials and not yet approved, though compounded topical forms exist in some markets.
Melatonin supplementation at night supports the natural nocturnal peak in thymulin and Ta1 that melatonin drives.
Mechanisms Of Action
Simple:
- The thymus secretes a family of peptide hormones that regulate immune function beyond the physical production of T cells.
- Thymosin fraction 5 (TF5) is a crude thymic extract containing more than 40 active peptides; its fractionation led to the isolation of all other thymic peptides.
- Thymosin alpha 1 (Ta1, 28 amino acids) activates dendritic cells via TLR3/4/9, promotes Th1 T cell differentiation, expands Tregs, enhances NK cell activity, and balances immune responses in both immunodeficiency and over-activation.
- Prothymosin alpha (ProTa, 109 amino acids) is the intracellular precursor to Ta1; extracellularly it acts as an alarmin that activates innate immunity via TLR4.
- Thymosin beta 4 (Tb4, 43 amino acids) sequesters G-actin intracellularly to regulate cytoskeletal dynamics; extracellularly it drives wound healing, angiogenesis, cardiac regeneration, and anti-inflammatory responses.
- Thymulin (nonapeptide) requires zinc for biological activity and acts on T cell precursors to promote differentiation; blood levels decline sharply with age and parallel the decline in immune competence.
- Thymopoietin (49 amino acids) drives T cell differentiation from bone marrow precursors and modulates mature T cell cytokine profiles via cGMP; its active site is thymopentin (TP5), a synthetic pentapeptide with the same activity.
- All thymic peptide levels decline with thymic involution, aging, chronic stress, and zinc deficiency; supplementation strategies can partially restore the hormonal environment these peptides normally provide.
Advanced:
- Ta1 TLR signaling specificity: Ta1 engages TLR3 on myeloid dendritic cells (recognizing dsRNA from virus-infected cells), TLR4 on macrophages (recognizing bacterial LPS and DAMPs), and TLR9 on plasmacytoid DCs and B cells (recognizing CpG DNA from bacteria and viruses). Each TLR engagement triggers distinct downstream cascades: TLR3-TRIF drives IRF3 and IFN-β (antiviral); TLR4-MyD88 drives NF-κB and pro-inflammatory cytokines (antibacterial innate); TLR9-MyD88 in pDCs drives IRF7 and massive IFN-α (antiviral surveillance). By activating all three pathways, Ta1 provides a broad-spectrum innate immune priming effect that subsequently shapes adaptive T and B cell responses.
- IDO pathway and immune tolerance: Ta1-activated myeloid DCs can simultaneously induce the enzyme indoleamine 2,3-dioxygenase (IDO), which catabolizes tryptophan to kynurenine in the local microenvironment. Kynurenine activates the aryl hydrocarbon receptor (AhR) on developing T cells, promoting Treg differentiation over Th17 differentiation. In inflammatory and autoimmune contexts, this IDO pathway modulates excessive immune activation toward a more tolerogenic phenotype, explaining how Ta1 can both activate immunity in immunodeficiency and moderate it in autoimmunity by operating on the same dendritic cell via the same activation event.
- Tb4-ILK-PI3K survival signaling: Extracellular Tb4 signals through the integrin-linked kinase (ILK) pathway after binding to its putative extracellular receptor. ILK phosphorylates AKT (protein kinase B), activating the PI3K-AKT-mTOR survival signaling axis. This promotes cardiomyocyte survival in hypoxia, keratinocyte survival at wound edges, and neuronal survival after ischemic injury. Simultaneously, Tb4-ILK signaling activates the PINCH and parvin scaffold proteins, coordinating actin cytoskeletal attachment to the extracellular matrix for cell migration. The anti-apoptotic and pro-migratory effects both emerge from the same ILK-mediated signal.
- Zinc-thymulin activity paradox in aging: In elderly individuals, both total thymulin peptide concentrations and zinc bioavailability decline, but they decline through different mechanisms. Thymulin peptide decline is driven by thymic epithelial cell loss (atrophy). Zinc bioavailability decline is driven by increased metallothionein (MT) expression in aging tissues, which sequesters zinc away from the extracellular compartment. Critically, high IL-6 (a feature of inflammaging) upregulates both glucocorticoid production and MT expression, creating a feed-forward loop: inflammation drives more zinc sequestration, which reduces thymulin activity, which impairs immune regulation, which sustains inflammation. This molecular loop explains why anti-inflammatory interventions (including zinc supplementation, which dilutes the zinc-binding capacity of MT) can partially break the immunosenescence spiral independent of any direct effect on the thymus itself.
Genetics
Legumain (LGMN) and Ta1 generation:
Legumain is the asparagine endopeptidase responsible for cleaving prothymosin alpha to generate thymosin alpha 1 at position 28.
LGMN polymorphisms that reduce legumain expression or activity would be expected to reduce Ta1 generation from ProTa, affecting circulating Ta1 levels.
LGMN activity is also pH-dependent (optimal at lysosomal pH), meaning cellular conditions that alter lysosomal function can affect Ta1 production even without genetic variation. R
TLR variants and Ta1 response:
Because Ta1 acts through TLR3, TLR4, and TLR9, the functional variants in these receptors (TLR4 Asp299Gly, TLR4 Thr399Ile, TLR9 rs352140) would be expected to modify Ta1's downstream effects.
Individuals with reduced-function TLR4 variants may have attenuated responses to Ta1's innate immune priming effects.
This connection has been established conceptually but not yet systematically in clinical pharmacogenomics studies.
Zinc transporter and thymulin activity:
Zinc bioavailability depends on the activity of zinc importers (ZIP family transporters, encoded by SLC39A genes) and zinc exporters (ZnT family, SLC30A genes), as well as metallothionein (MT1A, MT1B, MT2A) expression.
Genetic variants that increase metallothionein expression or reduce ZIP transporter efficiency would reduce intracellular zinc availability, impairing thymulin bioactivation.
Common MT1A variants have been associated with altered zinc homeostasis in aging.
VDR and thymic peptide rhythms:
Vitamin D receptor (VDR) signaling intersects with thymic peptide biology.
VDR activation promotes cathelicidin expression in thymus-adjacent innate immune cells and modulates the circadian coordination of thymic hormone secretion.
VDR polymorphisms (particularly FokI, BsmI, ApaI, TaqI) associated with altered VDR function may indirectly affect thymic peptide bioactivity by modifying the hormonal milieu in which thymic epithelial cells operate.
More Research
- Thymic regeneration strategies: Thymic involution is no longer considered an irreversible process.
Interleukin-22 (IL-22), keratinocyte growth factor (KGF), insulin-like growth factor 1 (IGF-1), and growth hormone have all demonstrated capacity to partially reverse thymic atrophy in animal models.
Sex steroid ablation (castration in animal models, LHRH agonist analogs in clinical trials) temporarily increases thymic size and output by removing sex hormone-mediated thymic involution.
FOXN1 gene therapy to restore thymic epithelial cell function is in early development.
The question of whether restoring thymic size also restores thymic peptide secretion is underexplored. R
- Ta1 in combination with immune checkpoint inhibitors: The therapeutic rationale is compelling.
Immune checkpoint inhibitors (anti-PD-1, anti-CTLA-4) release the brakes on T cell activation but cannot function if T cell numbers and quality are insufficient.
Ta1 restores the T cell pool, improves its functional state, and reduces exhaustion, potentially allowing checkpoint inhibitors to act on more and better-quality T cells.
Retrospective data from lung cancer is promising; prospective combination trial design is an active area. R
- ProTa as a cancer adjuvant and alarmin: ProTa's dual intracellular/extracellular biology positions it as a potential next-generation immunotherapy adjuvant.
Extracellular ProTa and its C-terminal decapeptide ProTa(100-109) activate TLR4 on DCs and macrophages, potentially providing adjuvant activity with a thymic peptide safety profile superior to standard TLR4 agonists used in vaccine adjuvant systems.
Clinical translation requires resolving the oncogenic risk of ProTa overexpression (its role as a proliferation marker in tumor cells) when used systemically. R
- Tb4 in post-MI cardiac regeneration: The combination of Tb4 and cardiac transcription factor reprogramming (Gata4/Mef2c/Tbx5) in a single therapeutic approach to post-infarction cardiac repair addresses multiple barriers simultaneously: Tb4 creates a permissive angiogenic and survival environment; the transcription factors convert resident cardiac fibroblasts to cardiomyocyte-like cells.
The scale of cardiac muscle replacement this approach might achieve in humans, and the safety of in vivo cardiac fibroblast reprogramming, remains to be determined in larger studies. R
- Thymopentin (TP5) and T cell exhaustion: The 2025 data showing TP5 reverses T cell exhaustion while preserving effector function opens an important mechanistic question: how does a pentapeptide acting through HLA-DR on APCs translate to rescue of exhausted T cells in the tumor microenvironment?
Epigenetic reprogramming of exhausted T cells typically requires complex interventions; if TP5 can reset exhaustion markers through thymic-dependent and thymic-independent mechanisms, this would represent a clinically practical approach to one of the major limitations of adoptive T cell therapy. R
- Microbiome and thymic peptide production: The gut microbiome influences thymic output through multiple routes: microbiome-derived metabolites (particularly short-chain fatty acids and secondary bile acids) affect VDR signaling, which in turn regulates antimicrobial peptides that shape the microbial environment that determines pathogen exposure.
Whether gut microbiome composition directly affects thymic epithelial cell function or thymic peptide secretion is not yet established, but the HPA-thymus-microbiome axis represents a plausible and underexplored research direction.
Jacob Gordon
INHC, FMT-C
Board Certified Health Coach
I spent years battling unexplained chronic illness before discovering biohacking, epigenetics, and functional medicine. Now I share that research at MyBioHack to help others find their own answers.
Book a ConsultationRelated Protocols & Supplements
Deep-dive chapters and recommended supplements for this topic
Quercetin
500mg 2x/day
Vitamin D3 + K2
5000 IU + 200mcg/day
Magnesium Glycinate
400mg at bedtime